 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Special Publication, No. 65, pp. 1-20
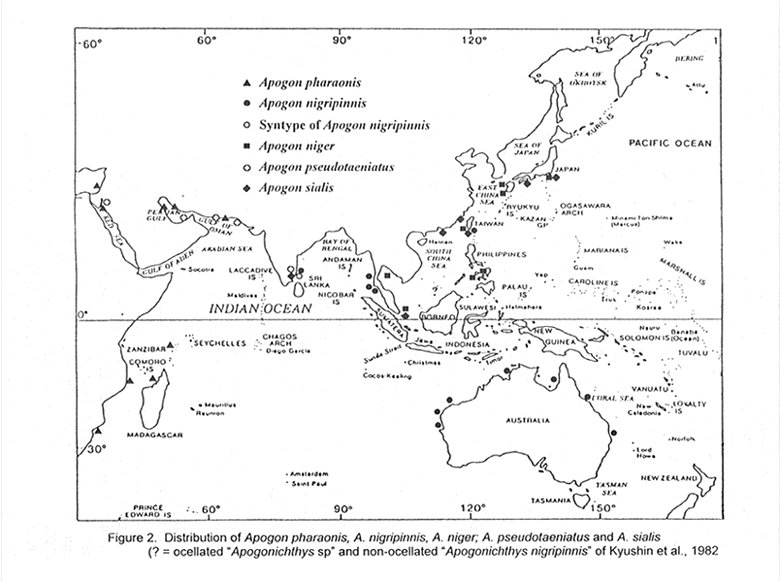
The Taxonomic Status of the Cardinalfish species Apogon Niger, A. Nigripinnis, A. Pharaonis, A. Sialis, and Related Species (Perciformes: Apogonidae) Offer Gon J. L. B. Smith Institute of Ichthyology November 2000 Code Number: fs00002 ABSTRACT Apogon niger Döderlein, 1883 of the western Pacific and Apogon nigripinnis Cuvier, 1828 (Apogon thurstoni Day, Amia nigripes Ogilby, and Amia atripes Ogilby are synonyms) of the eastern Indian Ocean and Australia are regarded as valid species distinct from Apogon pharaonis Bellotti, 1874 of the western Indian Ocean (Apogon suezii Sauvage, Amia ocellata Von Bonde, and Apogon duops Barnard are synonyms). Apogon nigripinnis differs from A. niger in having a large ocellus between the lateral line and the pectoral fin. A lectotype for A. niger is designated. A. pharaonis differs from both these species in having shorter pelvic fins, 16-21 instead of 15-18 gill-rakers, and 3 dark bars on the body. Apogon sialis (Jordan & Thompson, 1914), type locality Japan, is the senior synonym of Amia cathetogramma Tanaka, 1917, also described from Japan. Apogon pseudotaeniatus Gon, 1986 from the Red Sea, previously misidentified as A. bifasciatus Rüppell, is very similar in colour pattern to A. sialis; it differs in having a larger caudal spot, a dark instead of pale intestine, and in several morphometric characters. INTRODUCTION Apogon nigripinnis Cuvier has been reported from the Red Sea and western Indian Ocean (Regan, 1905; Smith, 1961; Dor, 1984; Gon, 1986a; Randall, 1995), Taiwan (Shen et al., 1993) and Australia (Gloerfelt-Tarp & Kailola, 1984; Sainsbury et al., 1984; Allen & Swainston 1988; Paxton et al., 1989). Day (1875), Steindachner & Döderlein (1883) and Weber & de Beaufort (1929) included Japan in the range. Yet, as is evident from colour illustrations in several recent publications (e.g. Gloerfelt-Tarp & Kailola, 1984; Randall, 1995; Kuiter, 1993; Kuiter & Kozawa, 1999), the fishes attributed to A. nigripinnis across this range exhibit different colour patterns. Allen, in Paxton et al., (1989) placed Apogon atripes Ogilby, originally described from Queensland, in the synonymy of A. nigripinnis; Kuiter (1992, 1993) disagreed and considered A. atripes a widespread West Pacific species similar to A. nigripinnis of the Indian Ocean coast of Australia. Recently, Kuiter in Kuiter & Kozawa (1999), revised his concept of A. atripes, restricting it to tropical Australia, with slight differences between east and west populations, and suggested that A. nigripinnis is a senior synonym of Apogon timorensis Bleeker. Kuiter & Kozawa (1999) raised Apogon cathetogrammus (Tanaka) from
obscurity. Although the colour pattern of this species is very similar to Apogon
pseudotaeniatus Gon from the Red Sea, these authors retained both as valid
species. The problems associated with Apogon nigripinnis, the rediscovery of A. sialis and A. cathetogrammus, and the apparent similarity of A. sialis to both of these species initiated this investigation. METHODS AND MATERIALS Measurements follow Gon (1993) and were taken to the nearest 0.5 mm. Ratios of body proportions were rounded to the nearest .05. Standard length (SL) is from the tip of upper jaw to the end of the hypural plate. Counts of pectoral-fin rays (which include the uppermost rudimentary ray) and lateral-line scales were made on both sides of the fish. The lateral-line count is of tubular scales to the end of the hypural plate + those extending onto caudal fin. A developed gill-raker is higher than the width of its base. The raker at the angle of the gill arch is included in the lower-limb count. When a raker is found on the ceratobranchial-hypobranchial joint it is included in the ceratobranchial count. The caudal spot is a midlateral dark spot posteriorly on caudal peduncle. The cheek mark is a narrow dark band from the ventral edge of the eye to the angle of the preopercular ridge. Specimens were borrowed from or examined at the following institutions: Australian Museum, Sydney (AMS); Natural History Museum, London (BMNH); Bernice P. Bishop Museum, Honolulu (BPBM); Field Museum of Natural History, Chicago (FMNH); Hebrew University, Jerusalem (HUJ); Museum of Comparative Zoology, Cambridge, USA (MCZ); Muséum National d'Histoire Naturelle, Paris (MNHN); National Science Museum, Tokyo (NSMT); Naturhistorisches Museum, Wien (NMW); J.L.B. Smith Institute of Ichthyology, Grahamstown (RUSI); Senckenberg Museum, Frankfurt (SMF); Western Australian Museum, Perth (WAM); United States National Museum of Natural History, Washington D.C. (USNM); Zoological Institute and Zoological Museum, University of Hamburg (ZMH); Zoological Museum Humboldt University, Berlin (ZMB); Department of Zoology, University Museum, University of Tokyo (ZUMT). MSNM is the Museo Civico di Storia Naturale, Milano; PMBC is Phuket Marine Biological Center, Thailand; QM is Queensland Museum, Brisbane and AFAQ is Amateur Fisheries Association of Queensland. The BMDP statistical software (Dixon et al. 1985) was used for statistical analysis of data. Normal probability plots were used to establish the approximate normality of data. Specimen data were grouped according to colour pattern (in alcohol) and geographical areas as follows: Western Indian Ocean, including the Red Sea and Arabian Gulf (ocellus present, a single dark bar under each dorsal fin [Apogon pharaonis] ); India and Australia (ocellus present, usually no bars on body [A. nigripinnis]); and western Pacific Ocean north of Australia (no ocellus, usually with narrow dark bars on body [A. niger]). The three groups were then compared for 8 meristic and 28 morphometric characters. Subgroup comparison was carried out to establish similarity between Red Sea and western Indian Ocean proper populations, between populations at the east and west coasts of Australia and between Japan and the western Pacific north of Australia. Large overlap of the range and mean excluded most of the characters from further analysis. The remaining characters (Table 1) were tested for significant differences between pairs of subgroups (T-test; P<0.001), and between the three main groups (Anova; P<0.001). Small subgroups (<20 specimens) and counts were also compared using a non-parametric test producing the same results. For the counts, a statistically significant difference of less than one unit in the mean of a character was considered unimportant at the species level (Table 1, pectoral rays and upper limb gill-rakers). Such differences may be useful at the population level. The fish length given in the caption of underwater photographs is an estimate of the total length. Otherwise, the lengths provided are the standard length. Apogon niger Döderlein, 1883 Apogon niger Döderlein, in Steindachner & Döderlein, 1883: 2 (Kochi, Japan; lectotype, NMW 35052). DIAGNOSIS: Dorsal fin VII + I,9; anal fin II,8, pectoral rays 15-17 (rarely 17); lateral-line scales 24 + 2-4 (rarely 26 or 28); predorsal scales 2 (rarely 1); gill-rakers 3-5 + 10-12 = 15-17, developed rakers 2-3 + 8-10, ceratobranchial rakers 7-8. Body depth 1.95-2.6 and head length 2.05-2.6 in SL; length of upper jaw 1.7-2.0 and of lower jaw 1.4-1.7 in head length; first dorsal-fin spine 1.5-3.5 in second spine; pelvic-fin length 2.6-3.5 in SL; pelvic-fin spine 1.4-2.2 in pelvic-fin length (including juveniles). Preopercle edge completely serrate, its ridge smooth, sometimes irregular; posttemporal serrate. Caudal fin truncate to slightly rounded. Colour in alcohol: body pale brown to brown, frequently with narrow dark brown bars; dark brown diffuse markings sometimes present on various parts of body; cheek mark usually present and, infrequently, another 1-2 faint narrow stripes radiating from posterior edge of eye; large dusky caudal spot, frequently overlain by dusky bar posteriorly on caudal peduncle; first dorsal and pelvic fins dark brown; second dorsal and anal fins dark brown, sometimes with narrow pale distal margin; caudal and pectoral fins pale; peritoneum with scattered small dark spots; intestine with varying amount of dusky streaks, rarely pale. Ontogenetically, in juveniles, dark pigment seems to spread from front of body backwards; pale second dorsal and anal fins darken from base distally (Fig. 1A&B); several dark bars present in a 21.3 mm SL fish (Fig. 1B). Colour in life (photographs in Kuiter & Kozawa ,1999): body reddish to purplish dark brown, frequently with blackish narrow bars varying in intensity; narrow cheek mark present but sometimes faint; dark caudal spot overlain by dark bar posteriorly on caudal peduncle; dorsal, anal and pelvic fins dark brown to black; caudal fin pale to yellow. Figure 1. Colour patterns of juveniles of Apogon niger, NSMT 53595, 13 mm SL (A) and 21.3 mm SL (B); A. nigripinnis, AMS 17664-001, 19.1 mm SL (C); and A. pharaonis, HUJ 11803, 21 mm SL (D). DISTRIBUTION: Apogon niger is known from the Gulf of Thailand, Singapore, Philippines, Taiwan, and Japan (Fig. 2). REMARKS: Japanese and western Pacific (north of Australia) specimens differed significantly in the length of the third dorsal-fin spine, depth of caudal peduncle and pelvic fin/spine ratio, but these differences were not significant (P = 0.02, 0.01, 0.06, respectively) when juveniles were removed from the Japanese sample, as there were no juveniles in the western Pacific sample. There were no significant differences between samples from the east and west coasts of Australia. Comparing Apogon niger, nigripinnis and pharaonis: the caudal peduncle depth of Apogon niger and the peduncle length of A. nigripinnis were significantly different from the other two species, respectively. However, in both characters, the difference in the mean between species is less than 2% SL, and the range is completely overlapping, or nearly so (Table 1). These characters are therefore of little taxonomic use. The length of the pelvic fin of A. pharaonis was significantly different from the other two species, but this character was not significantly different (P = 0.8) between niger and nigripinnis. Although the length of the pelvic-fin spine is not significantly different between one species and another (P = 0.02), the adult pelvic fin/spine ratio is a good diagnostic character due to the shorter pelvic fin of pharaonis. This ratio is 1.45-1.75 in adult pharaonis and 1.7-2.2 in adult niger and nigripinnis. The preceding analysis of morphometric and colour characters shows that Apogon pharaonis is distinctly different from both of the other species (Tables 1-3; see also Remarks for A. pharaonis below). Apart from the ocellus, A. niger and nigripinnis are the same, and the taxonomic status of these two taxa is problematic. Because there is no knowledge of the biology of the two forms, their status may be inferred by finding intermediate colour forms. The fish depicted in Kyushin et al. (1982: 191) may be an intermediate form, but the unusual colour pattern on its caudal fin calls for verification with actual specimens. In the absence of clearly intermediate forms and in view of the stability of the ocellus character in Australia (always present) and the western Pacific north of Australia (always absent), including very small juveniles, two species are recognised here. However, a hypothesis of two allopatric species provides no explanation for the presence of the ocellated fish in Taiwan and the non-ocellated type of A. nigripinnis in India (see also Remarks for A. nigripinnis below). The use of molecular taxonomic methods to determine the level of genetic differentiation between the two forms may help resolve this taxonomic problem. Two other species of the Apogon nigripinnis species group, i.e.A. pharaonis Bellotti and A. taeniatus Cuvier, have an ocellus similar to that of A. nigripinnis. In life, the ocellus of these species may be incomplete, present on one side only, or absent altogether. In addition, in A. pharaonis the ocellus always shows up after death and preservation, but in A. taeniatus adults without an ocellus frequently remain so after death (Gon & Randall, in press; Smith, 1961: 394, 396). These observations suggest that these two species can control the display of the ocellus. The fact that specimens of A. niger never show an ocellus lends support to the hypothesis that the Australian and western Pacific forms are different species. A study of the variation and function of the ocellus in the A. nigripinnis species group is necessary to confirm this hypothesis. The name Apogon niger Döderlein was first published by Steindachner & Döderlein (1883: 2) in the synonymy of A. nigripinnis, and as such it is an unavailable name. However, as pointed out by Eschmeyer et al. (1998), usage may render it available. The International Code of Zoological Nomenclature (Article 11.6) states that such a name becomes available if it was treated as available and adopted as a name for a taxon before 1961. Jordan & Snyder (1901: 895, fig. 2) treated A. niger Döderlein as available; these authors, Smith & Pope (1906: 468), Jordan et al. (1913: 139, fig. 96), Jordan & Hubbs (1925: 230), and Kamohara (1955: pl. 28), applied this name to the Japanese species described by Steindachner & Döderlein (1883). Therefore, this name is here regarded as an available name. Steindachner & Döderlein (1883) refer to 3 specimens of which the largest is "...somewhat over 9 cm long." The Natural History Museum of Vienna has 3 specimens labelled "Apogon nigripinnis" (NMW 35052-54), which were collected by Döderlein and donated by Steindachner. The locality on the jar labels is Tokyo, but all 3 fish have small pencilled labels in their mouths with the name Kochi. A comparison of the writing on these labels with the handwriting of Steindachner and of Döderlein in documents at the NMW archives revealed that Döderlein was probably the one who had written them (H. Wellendorf, NMW, pers. comm.). These 3 fish agree with the description of A. nigripinnis in Steindachner & Döderlein (1883) and are here regarded as Döderlein's original specimens of A. niger. The largest specimen (NMW 35052, 72.6 mm SL, 93.5 mm TL) is designated here as the lectotype of Apogon niger. Three other specimens labelled Apogon nigripinnis (NMW 35049-51) and re-identified here as Apogon ellioti Day, are probably a part of the series of 4 specimens (NMW 34883 is the fourth) used by Steindachner & Döderlein (1883) for the description of A. ellioti. Oral incubation of eggs by male A. niger was reported by Hayashi (1984). MATERIAL EXAMINED: Thailand: Gulf of Thailand, Go Sichang, 4: 42.7-56.4 mm. Singapore: Sultan Shoal, BMNH 1984.1.13.65-71, 6: 45.7-67.5 mm. Taiwan: RUSI 38435, 44.8 mm. Philippines: Iloilo, Panay Island, USNM 358065, 55.3 mm; Samar Sea, Carigara Bay, USNM 358066, 72.0 mm; SE Malangaban Island, USNM 276704, 10: 59.0-70.6 mm; USNM 276708, 2: 64.5-74.0 mm. Japan: Nagasaki, USNM 49925, 7: 68.9-77.1 mm; NSMT-P 53134, 23: 11.8-30.6 mm; Tachibana Bay, NSMT-P 53595, 34: 13.3-39.7 mm; Kagoshima, USNM 59628, 62.1 mm; Shikoku, Susaki, USNM 59629, 68.6 mm; Kochi, NMW 35052, 72.6 mm (male, lectotype of Apogon niger Döderlein); NMW 35053, 68.7 mm and NMW 35054, 60.6 mm (paralectotypes of Apogon niger Döderlein); Suruga Bay, USNM 71361, 2: 73.6-78.2 mm; Tokyo Bay, BMNH 1891.5.26.4, 67.2 mm. Apogon nigripinnis Cuvier, 1828 Apogon nigripinnis Cuvier, 1828: 152 (type localities, Pondicherry,
India and Java, Indonesia; Indian syntype, MNHN 8694; Indonesian syntype lost). DIAGNOSIS: Dorsal fin VII + I,9; anal fin II,8, pectoral rays 15-17 (rarely 17); lateral-line scales 24 + 3-4 (rarely 28); predorsal scales 2; gill-rakers 4-5 + 11-13 = 15-18 (1 of 32 with 18); developed rakers 2 + 8-10; ceratobranchial rakers 7-8. Body depth 1.9-2.4 and head length 2.0-2.5 in SL; length of upper jaw 1.7-2.0 and of lower jaw 1.4-1.7 in head length; first dorsal-fin spine 1.7-2.85 in second spine; pelvic-fin length 2.5-3.7 in SL; pelvic-fin spine 1.5-2.3 in pelvic-fin length. Preopercle edge completely serrate, its ridge smooth; posttemporal serrate. Caudal fin truncate to slightly rounded. Colour in alcohol, body pale brown to brown, sometimes with several narrow dark brown bars; large dark brown ocellus anteriorly between lateral line and pectoral fin; cheek mark usually present; occasionally, another 1-2 faint narrow stripes radiating from posterior edge of eye, large dusky caudal spot, frequently overlain by dusky bar posteriorly on caudal peduncle; first dorsal and pelvic fins dark brown; second dorsal and anal fins dark brown, the former usually with pale distal margin of varying width and latter fin more rarely so; caudal and pectoral fins pale; peritoneum with scattered small dark spots; intestine with varying amount of dusky streaks, rarely pale. Ontogentically, dark pigment seems to spread from front of body backwards; fish smaller than about 35 mm TL have relatively pale caudal peduncle (except caudal spot); the ocellus develops in fish smaller than 20 mm as a dark spot (Fig. 1C), the pale ring around it appearing at about 30 mm; pale second dorsal and anal fins darken from base distally. Colour in life (photographs in Sainsbury et al., 1984; Kuiter, 1992,1993, as Apogon atripes, and Kuiter & Kozawa, 1999, as Apogon atripes): body reddish to purplish dark brown, sometimes with narrow bars varying in intensity; narrow cheek mark present but sometimes faint; large dark brown to black, yellow-edged ocellus anteriorly between lateral line and pectoral fin; dark caudal spot, overlain by dark bar posteriorly on caudal peduncle; first dorsal, anal and pelvic fins dark brown to black; second dorsal and anal fins dark brown, the former with pale to yellowish distal edge varying in width; caudal fin pale to yellow. DISTRIBUTION: Apogon nigripinnis is known from the east coast of India, Andaman Sea and Australia (Fig. 2). REMARKS: Apogon nigripinnis is distinguished from A. niger in having a large dark ocellus above the pectoral fin and a pale distal edge of the second dorsal fin (see also Remarks for A. niger above). All the Australian specimens (smallest is 19.1 mm SL) and photographs of Australian fish examined for this study have the midlateral ocellus and usually no narrow dark bars on the body. However, the underwater photograph of an adult Apogon atripes in Kuiter (1993: 155) shows an ocellus and at least 7 dark bars, as do several WAM specimens from Western Australia and Northern Territory. Ocellated specimens from north of Australia are rare in collections, and none were examined in this study. An ocellated fish was reported from Taiwan (Shao & Chen, 1986: 90, fig. 34, as Apogon niger; Shen et al., 1993, pl. 83, fig. 4, as Apogon nigripinnis). Kyushin et al. (1982: 191) illustrated a fish (as Apogonichthys sp.) from the South China Sea with a faint ocellus and a gill-raker count that agree with nigripinnis, but the dark patches on the caudal fin do not. Kyushin et al. (1982: 190) also illustrated a non-ocellated fish as "Apogonichthys nigripinnis," but this fish has a dark caudal fin and is probably a different species. The non-ocellated form of Shen et al. (1993, pl. 83, fig. 3, as niger) is the same as in Japan. Kuiter & Kozawa (1999) present underwater photographs of yellowish individuals, identified as A. niger, of which one has a smaller black spot instead of the ocellus. The photograph of a yellowish 'niger' by Abe (1996) depicts a similar spot. However, the identification of the yellowish form as A. niger, as defined above, needs to be verified. Amia atripes Ogilby is included in the synonymy of A. nigripinnis even though his holotype is lost. The original illustration and description provide enough diagnostic characters (e.g. ocellus, cheek mark, long dark pelvics, dark second dorsal with pale distal edge, and pale caudal) to identify his specimen as A. nigripinnis. Kuiter in Kuiter & Kozawa (1999) identified ocellated fish from tropical Australia as Apogon atripes and postulated that A. nigripinnis may be a senior synonym of Apogon timorensis Bleeker, 1854. This is unlikely because A. timorensis has fewer developed gill-rakers (1 + 6-8), more predorsal scales (3), and no caudal spot. Kuiter may have been misled by the figure and description of A. nigripinnis in Day (1875) which portray no ocellus and list several characters that can be interpreted either way. As implied above, Day apparently confused A. nigripinnis with another species, possibly A. timorensis. The extant syntype of A. nigripinnis (MNHN 8694) from India has no trace of an ocellus, and Cuvier (1828) made no reference to one in his brief description of this species. The presence of the non-ocellated form (= A. niger) in Indian waters could not be verified because Indian specimens are very rare in collections. Such presence is unlikely in view of the apparent absence of A. niger in the Andaman Sea off the west coast of Thailand (U. Satapoomin, PMBC, pers. comm.). Day's (1875) description of a non-ocellated A. nigripinnis from Madras, stated to be "identical" with Cuvier's type, is also questionable. His colour description does not agree with any of the specimens examined for this study, ocellated or not, including Cuvier's type. Moreover, one of his specimens from Madras (BMNH 1889.2.1.2891), labelled Apogon nigripinnis, clearly has the ocellus. In the absence of direct evidence either way, applying the name A. nigripinnis to the ocellated species serves to maintain nomenclatural stability. MATERIAL EXAMINED: India: Madras, BMNH 1889.2.1.2891, 48.4 mm (collected by Day); BMNH 1889.8.7.2, 54.2 mm (holotype of Apogon thurstoni); Pondicherry, MNHN 8694, 62.9 mm (syntype of Apogon nigripinnis). Thailand: Ranong, PMBC 10360, 52.0 mm; Phuket Island, PMBC 14633, 47.3 mm; Lanta Yai Island, PMBC 14632, 47.9 mm. Australia: Western Australia, NW shelf, WAM P.28750-001, 2: 54.2-60.6 mm; Exmouth Gulf, WAM P.25508-014, 53.5 mm; WAM P.27224-007, 2: 53.0-58.3 mm; Shark Bay, WAM P.13755-001, 63.4 mm. Northern Territory, Darwin, WAM P.14472-001, 61.2 mm. Queensland, Gulf of Carpentaria, WAM P.12784-001, 68.5 mm; Cape York, AMS I.2077154-022, 42.6 mm; Lizard Island area, AMS I.20754-025, 64.0 mm; Magnetic Island, MCZ 38497, 57.2 mm; Bowen, AMS E.2667, 67.0 mm; Double Bay, AMS E.2862, 66.9 mm; Port Clinton, AMS I.34397-008, 35.2 mm; NW of Hervey Bay, AMS E.2910, 2: 73.0-73.6 mm; E of Mooloolaba, AMS I.38246-001, 47.8 mm; Moreton Bay, AMS I.19577-002, 26.8 mm; WAM P.28826-001, 31.0 mm. New South Wales, Hawkesbury River, AMS I.34639-001, 51.5 mm; Sydney, AMS I.16194-001, 32.9 mm; AMS I.17326-001, 46.8 mm; AMS I.17664-001, 3: 19.1-63.4 mm; AMS I.17748-001, 25.0 mm; AMS I.17660-003, 2: 35.5-49.5 mm; AMS I.19484-001, 42.3 mm; AMS I.23881-005, 49.6 mm. Apogon pharaonis Bellotti, 1874 Apogon pharaonis Bellotti, 1874: 264 (Suez, Red Sea; holotype MSNM,
destroyed in 1943). DIAGNOSIS: Dorsal fin VII + I,9; anal fin II,8, pectoral rays 14-17; lateral-line scales 24 + 3; predorsal scales 1-3 (rarely 1 or 3); gill-rakers 4-6 + 12-15 = 16-21; developed rakers 2-3 + 9-12; ceratobranchial rakers 8-9. Body depth 2.0-2.5 and head length 2.1-2.5 in SL; length of upper jaw 1.8-2.2 and of lower jaw 1.6-1.8 in head length; first dorsal-fin spine 1.9-3.0 in second spine; pelvic-fin length 3.1-4.4 in SL; pelvic-fin spine 1.45-1.75 in pelvic-fin length. Posterior edge of preopercle serrate; serrae on anterior part of ventral edge minute, sometimes absent; preopercle ridge smooth; posttemporal serrate. Caudal fin truncate to slightly emarginate. Colour in alcohol: body pale brown to brown with 3 dark brown bars; first bar from anterior part of first dorsal base, containing large dark brown spot ringed in white above middle of pectoral fin (ocellus sometimes faint or absent on one side); second bar between bases of second dorsal and anal fins, and extending vertically onto both fins to about tip of their spine (2nd spine in anal fin); third bar at caudal-fin base, masking slightly wider caudal spot; faint bars frequently present between main 3 bars; narrow oblique cheek mark present; leading edge of first dorsal fin dark brown, the remainder dusky; second dorsal and anal fins pale to dusky (except dark brown extension of second bar); caudal fin pale to dusky; pelvic fins dusky to brown on proximal part, becoming dark brown distally; peritoneum with dark spots of various sizes; intestine pale. Juvenile pattern similar to adult, but bars tend to be of more intense colour. Colour of freshly collected specimens: similar to preserved pattern; generally bronze dorsally to silvery ventrally and with bars usually dark brown to black; first dark bar joins dark leading edge of first dorsal fin; second dark bar extends at least to middle of second dorsal and anal fins; no faint stripes on body. DISTRIBUTION: A. pharaonis is known from the Red Sea, Persian Gulf, Pakistan, Iran, Zanzibar, Mozambique, South Africa (north of Durban), Seychelles and Madagascar (Fig. 2). It is common in shore on silty reefs, seagrass beds, and in mangrove areas. REMARKS: Apart from its different colour pattern, Apogon pharaonis is distinguished from A. nigripinnis and A. niger in having more gill-rakers (Table 2), and a shorter pelvic fin (2.6-3.5 in SL in the other two species). Although the pelvic-fin spine has a similar size in all three species (Table 1), the shorter fin of adult pharaonis makes its fin/spine ratio distinctly different (1.7-2.3 in adult nigripinnis and niger). Juvenile pharaonis (Fig. 1D, smallest examined is 21.0 mm SL) have the same colour pattern as the adult. The photograph of a 30 mm TL juvenile nigripinnis in Kuiter & Kozawa (1999, as atripes) depicts a fish with a dark body, pale caudal peduncle with a large caudal spot overlain by a dark bar, completely dark first dorsal and pelvic fins and pale second dorsal and anal fins. The only common features in the juveniles of both species are the ocellus, the narrow cheek mark and the pale caudal fin. The differences between the juveniles are also evident in preserved fish (Fig. 1, C & D). Plate 1. A) Apogon niger, 10 cm, Ehime Japan (R.H. Kuiter; B) Apogon nigripinnis, Sydney Harbour, Australia (R.H. Kuiter); C) Apogon pharaonis, 75 mm, Persian Gulf, Saudi Arabia (J.E. Randall); D) Apogon sialis, 9 cm, Japan (J.E. Randall); E) Apogon sialis, 14 cm, Japan (R.H. Kuiter). F) Juvenile Apogon pseudotaeniatus, 5 cm, Gulf of Aqaba, Red Sea (O. Gon). Red Sea and western Indian Ocean specimens differed significantly in four characters (Table 4). Red Sea fish have a slightly shallower body, and slightly shorter first dorsal spine and pelvic fin; the differences in the mean of these characters amount to 3.3, 0.9 and 3.0 percent SL, respectively. In addition, the mean number of pectoral rays in Red Sea fish is 0.8 ray lower than in western Indian Ocean fish. The body depth and pelvic fin differences are due to the high number (7 of 13) of small juveniles in the Red Sea sample. Juveniles are more slender and the pelvic fin is shorter than in adults. These differences may not show in a comparison of larger, homogeneous samples. Removing Red Sea fish from the western Indian Ocean sample had no effect on the differences of the three species. In the comparison of the three main geographical areas, the small difference in mean body depth of Apogon pharaonis and A. nigripinnis was not significant (P = 0.2). Playfair & Günther (1867: 20) and Regan (1905: 330), though merely listings, are probably the earliest reports of Apogon nigripinnis from the western Indian Ocean. Playfair & Günther followed Günther's (1859) concept of A. nigripinnis, but Day (1881) referred their fish, as well as Günther's fish, to his Apogon ellioti. Indeed, their Zanzibar specimen (BMNH 1867.3.7.636) is Apogon hungi Fourmanoir & Nhu-Nhung, 1965, a close relative of A. ellioti (Gon, 1996). Another Playfair fish (BMNH 1869.1.79.41) from Zanzibar, labelled just Apogon, is pharaonis. Examination of Regan's (1905) specimens from the Persian Gulf (BMNH 1899.5.8.23) and the Arabian Sea (BMNH 1901.1.29.5-14) revealed the same ocellated form later described by Smith (1961: 395, pl. 48A) as Apogonichthyoides nigripinnis (Cuvier). Apparently no other workers in the western Indian Ocean associated this form with the name A. nigripinnis Cuvier prior to Smith (1961). It is therefore not surprising that during that period of time the ocellated form of the western Indian Ocean was described several times (see list of synonyms above). Barnard (1927: 522) wrote that the holotype of Amia ocellata von Bonde was in the collection of the Fisheries & Marine Biological Survey (later replaced by the Sea Fisheries Research Institute) of South Africa. This fish was part of a large collection made during the 1920s by the S.S. Pickle in South African waters. Unfortunately, this collection was allowed to deteriorate to the point of destruction (Winterbottom ,1974; Gon & Skelton, 1997). Several type specimens from this collection were rescued and are found at the J.L.B. Smith Institute of Ichthyology, the South African Museum, Cape Town, and the Natural History Museum, London. The holotype of von Bonde's species was not found in any of these institutions and is therefore presumed lost. Authors of recent papers on western Indian Ocean fishes followed Smith (1961) in using A. nigripinnis Cuvier for this species. Smith (1961), however, expressed some reservations when he used A. nigripinnis because he could not reconcile the colour pattern of his specimens with Cuvier's type. He was also not sure about the status of A. thurstoni Day used by previous authors for Red Sea and Arabian Gulf specimens of pharaonis (see Blegvad & Loppenthin, 1944 and synonymy for A. nigripinnis in Dor, 1984). Fraser (2000) compared ocellated specimens from southern Africa with the holotype of nigripinnis Cuvier and concluded that they are that species. However, his analysis was limited to demonstrating that Smith's demotion of Apogon gardineri Regan to a junior synonym of nigripinnis was incorrect. MATERIAL EXAMINED: Mediterranean: Israel, Akko, HUJ 13584, 75.4 mm. Red Sea: Gulf of Suez, Suez, MNHN A-5137, 50.2 mm (holotype of suezii); Ras Misalla, HUJ 11803, 6: 21.0-34.9 mm; HUJ 11810, 53.6 mm; east of Deversoir, HUJ 5767, 55.8 mm; Ras es Sudr, HUJ 5764, 4: 31.1-54.8 mm. Zanzibar: BMNH 1869.1.79.41, 50.6 mm. Mozambique: Ibo, RUSI 5808, 3: 54.3-65.1 mm; Quilaluia, RUSI 5642, 3: 45.5-63.4 mm; Inhaca Island, RUSI 56383, 14: 42.4-63.7 mm. Seychelles: Aldabra, RUSI 3153, 2: 48.6-54.2 mm. Madagascar: ZMH 14265, 65.0 mm. Kuwait: Kuwait City, MCZ 59291, 2: 64.9-66.6 mm. Iran: Bushire, BMNH 1899.5.8.23, 62.5 mm; Charbar, BMNH 1901.1.29.5-14, 6: 26.0-47.8 mm. Apogon sialis (Jordan & Thompson, 1914) Amia sialis Jordan & Thompson, 1914: 246, pl. 28, fig. 2 (Suruga
Bay, Japan; holotype, FMNH 57084). DIAGNOSIS: Dorsal fin VII + I,9; anal fin II,8, pectoral rays 14-15 (rarely 14); lateral-line scales 24 + 3-5; predorsal scales 3; gill-rakers 5 + 13-14; developed gill-rakers 2 + 8-10. Body depth 2.2-2.5 and head length 2.3-2.5 in SL; length of upper jaw 2.0-2.3 and of lower jaw 1.75-1.9 in head length; third dorsal-fin spine 1.5-2.0 in head length; pelvic-fin length 3.2-4.0 in SL; pelvic-fin spine 1.4-1.7 in pelvic-fin length. Posterior edge of preopercle with small serrae; serrae on anterior half of ventral edge minute or absent; preopercle ridge smooth; posttemporal serrate; caudal fin emarginate. Colour in alcohol: pale brown to brown with two dark brown bars, one under anterior base of each dorsal fin; bars joining dark leading edge of both dorsal fins; dark narrow cheek mark sometimes present; small dark caudal spot 3.5-5.2 in caudal peduncle depth; first dorsal fin behind dark leading edge brown; second dorsal, anal and caudal fins dusky to brown, becoming paler distally; pelvic fins brown to dark brown; peritoneum pale with small blackish spots; intestine pale. Colour in life (underwater photographs in Kuiter & Kozawa 1999): body brownish to greenish grey anteriorly, shading to pale grey posteriorly; dark bars intense dark brown, reaching ventrally to level of lower pectoral-fin base or slightly lower; bars joining equally dark leading edge of both dorsal fins; remaining part of first dorsal fin white and that of second dorsal fin translucent; pelvic fins dark brown with white leading edge; distinct dark, small caudal spot. DISTRIBUTION: This species is known from the east coast of India, Singapore, China, Taiwan and Japan (Fig. 2). The distributional records of western Pacific A. pseudotaeniatus extracted from the literature by Gon (1986b: map 1) are likely to be sialis. Nevertheless, they are excluded from Fig. 2 as they need to be verified. REMARKS: Jordan & Thompson (1914) described the colour of Apogon sialis as "... nearly uniform, pattern lacking, save for an indistinct, small caudal spot; ventrals and spinous dorsal dark...", counted 5 + 13 gill-rakers and related their species to Apogon niger. However, unlike niger their holotype has traces of a dark bar under each dorsal fin (viewed under magnification), and no trace of a dark bar posteriorly on caudal peduncle or a narrow cheek mark (M. Westneat, FMNH, pers. comm.) In addition, the length of the upper jaw of the holotype (2.2 in head length) is distinctly shorter than in niger (1.7-2.0 in head length). These observations and the small caudal spot of the holotype agree with Apogon pseudotaeniatus Gon and Apogon cathetogrammus (Tanaka). Comparison with these two species led to the conclusion that sialis is identical to cathetogrammus and to West Pacific specimens of pseudotaeniatus. The name Amia sialis probably went into obscurity because it could not be associated with any of the cardinalfish species inhabiting Japanese waters. Amia cathetogramma suffered the same fate because authors identified the fish described by Tanaka (1917) either as Apogon taeniatus Cuvier, 1828 or as Apogon bifasciatus Rüppell, 1838 (see list of misidentifications in Gon, 1986b: 11). Gon (1986b) was unaware of these two forgotten Japanese species when he described Apogon pseudotaeniatus (Plate I F) and his paratypes include three West Pacific specimens (ZMB 53 and ZMH 14340, 14342) reidentified here as A. sialis. Although the colour pattern of Apogon sialis is virtually identical to that of A. pseudotaeniatus, several differences warrant keeping the latter as a valid species (Tables 2 & 5). The most noticeable difference is the higher first dorsal fin of sialis (third dorsal spine 2.0-2.45 in head length in pseudotaeniatus). This character was also noted and illustrated by Day (1875, p. 59, pl. 8, fig. 4 [A. sialis, identified as Apogon taeniatus] and p. 62, pl. 16, fig. 9 [A. pseudotaeniatus, identified as Apogon bifasciatus]; see Gon and Randall [in press] for a detailed treatment of A. taeniatus and pseudotaeniatus). Other differences between A. sialis and pseudotaeniatus that show little or no overlap are the colour of the intestine (dark in pseudotaeniatus), lengths of upper and lower jaw, lengths of second dorsal and pelvic-fin spines, and the caudal spot diameter. These differences are not age- or size-related, because the material of both species consist of adults of a similar size range (Table 5), and are therefore more reliable for separating the species. Nevertheless, the validity of pseudotaeniatus should be tested by studying more specimens of both species. MATERIAL EXAMINED: Japan: Suruga Bay, FMNH 57084, 90.4 mm, holotype Amia sialis; Tanabe, ZUMT 55054, 86.9 mm, holotype of Amia cathetogramma. Singapore: ZMH 14340, 69 mm, paratype of Apogon pseudotaeniatus. China: BMNH uncatalogued, 88.5 mm; ZMB 53, 95.8 mm, paratype of Apogon pseudotaeniatus; Hong Kong, BMNH 1939.1.17.40, 96.7 mm; Fukien Province, ZMH 14342, 61.4 mm, paratype of Apogon pseudotaeniatus. OTHER MATERIAL: Apogon pseudotaeniatus: Red Sea: Gulf of Aqaba: Egypt, off Nelson's Village, BPBM 27395, 86.8 mm, holotype; and BPBM 30551, 86.5 mm, paratype; Jordan, Aqaba, SMF 15948, 60.6 mm, paratype; Gulf of Suez, HUJ 9085, 87.2 mm. Persian Gulf: BMNH 1900.5.9.25, 56.9 mm. Iran: Chabar, BMNH 1901.1.29.3, 89.5 mm. Pakistan: Karachi, BMNH 1898.11.18.2-5, 3: 72.3-81.7 mm; BMNH 11898.12..24.5, 109.7 mm; BMNH 1901.30.86-87, 2: 71.7-102.2 mm.; BMNH 1903.5.14.3, 104.5 mm. ACKNOWLEDGEMENTS Thanks are due to M. McGrouther (AMS), O. Crimmen (BMNH), A. Suzumoto (BPBM), D. Golani (HUJ), A. Bentley and D. Sandi (RUSI), K. Hartel (MCZ), M. Bauchot, M. Dessouter, G. Duhamel and P. Pruvost (MNHN), K. Matsuura and G. Shinohara (NSMT), W. Klausewitz (SMF), S. Morisson (WAM), S. Jewett and D.G. Smith (USNM), H.-J. Paepke (ZMB), G. Schulze and H.Wilkens (ZMH), K. Sakamoto (ZUMT) for loans of specimens and use of museum facilities. T.H. Fraser (Mote Marine Laboratory, Florida), B. Chernoff, A. M. Rogers, and M. Westneat (FMNH) kindly provided data for the holotype of Apogon sialis. W.N. Eschmeyer provided advice on nomenclature. I am very grateful to H. Wellendorf (NMW) for his efforts in locating and identifying the syntypes of Apogon niger. E. Heemstra (RUSI) prepared the artwork. R.H. Kuiter and J.E. Randall kindly provided colour photographs. T.H. Fraser and J.E. Randall provided useful comments on the manuscript. Table 1. Measurements (as % SL) and counts of Apogon niger, A. nigripinnis and A. pharaonis. The range is followed (on parentheses) by the mean, standard deviation and sample size (n). The asterisk indicates a statistcally significant difference of the mean (P<0.001)
Table 2. Frequency distributions of gill-raker counts for Apogon pharaonis, A. nigripinnis, A. niger, A. sialis and A. pseudotaeniatus.
1 - holotype of A. suezii;
Table 3. Frequency distributions of left pectoral-fin ray counts in Apogon niger, A. nigripinnis and A. pharaonis.
1 - holotype of A. suezii;
Table 4. Comparison of Apogon pharaonis from the Red Sea and western Indian Ocean. The range is followed (in parentheses) by the mean, standard deviation and sample size (n); measurements are in % SL.
Table 5. Ranges of measurements (in % SL) of 5 A. sialis, 61.4 - 96.7 mm and 4 A. pseudotaeniatus, 56.9 - 110 mm.
LITERATURE CITED
The following images related to this document are available:Photo images[fs00002c.jpg]Line drawing images[fs00002b.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}