 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Special Publication No.60, May 1998 A LATE CRETACEOUS (MAASTRICHTIAN) GALAXIID FISH FROM SOUTH AFRICA
Code Number:FS98001
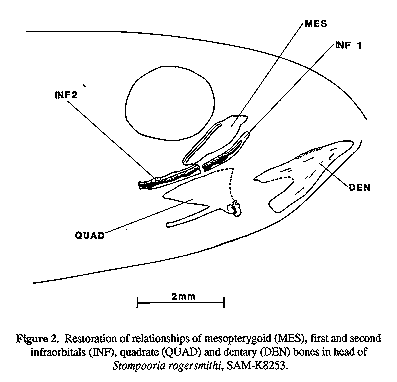
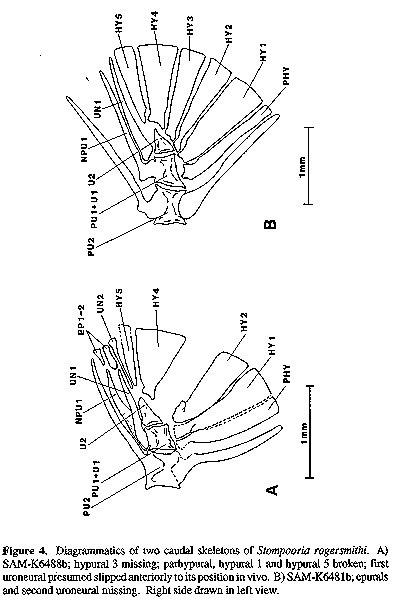
A new teleost fish, Stompooria rogersmithi, is described from deposits of a Late Cretaceous crater lake in the Karoo, South Africa. The fossils represent the earliest known species of the recently redefined family Galaxiidae. Distinguishing features include the apposition of the dorsal and anal fins, 41-42 vertebrae, all neural arches anterior to the dorsal fin origin fused to the centre, presence of epineurals, the opercle reduced above the level of the hyomandibular articulation, the posttemporal and supracleithrum without a connection to the seismosensory head canals, 18 principal caudal fin rays, two epurals, five hypurals, no dorsal outgrowth on the uroneural, the neural spines of the second and third preural centra similar in size, and absence of a stegural, a third uroneural, a mesocoracoid, a supramaxilla, and teeth on the maxilla and palatine bones. Semper aliquid novi Africa affert (Pliny the EIder, 23-79 A.D.) In January 1984, a De Beer's Consolidated Mines drilling team was prospecting for kimberlite pipes (diamond-bearing ore) near Marydale, Prieska District, Northern Cape Province, South Africa. A 12 cm diameter borehole sunk on the farm Stompoor yielded a core extending from 42 to 168 m below the surface near the center of the sediment-filled remains of a crater lake produced by maar-type volcanism. The core was dated palynologically to about 70 Ma, or the lower Maastrichtian (Smith, 1986; as Senonian). Sections through the ancient lake core were found to be fossiliferous, containing ostracods, gastropods, mussels, insects, a frog (either genus Eoxenopoides or Thoraciliacus; Van Dijk, 1985), a possible bird bone, and a previously unknown salmoniform fish, herein described. Smith (1986, 1995) described the Late Cretaceous Karoo crater lake habitat and included an artist's rendition of it, now part of a public display at the South African Museum, Cape Town (SAM). Smith (1988) reported on the geology of the 76 m of resin encased core that was preserved for study and mentioned again some of the fossils. He concluded a time span of some 220,000 years was represented by the 76 m of sediment accumulation. At first the Stompoor fishes were thought to be a primitive cyprinoid, but on closer examination were diagnosable within Johnson and Patterson's (1996) redefined family Galaxiidae. This was possible on the basis of determining that the species was a euteleost without a Weberian apparatus while failing into Patterson' s (1970:291 ) first caudal skeleton group (without a stegurai: aiepocephaioids, argentinoids, gaiaxioids and ostariophysans). Further characters that corroborate its placement are given in the systematic section below. MATERIALS AND METHODS Core samples were stablized in the field or laboratory with plaster moldings and/or glyptal painting. Bones were found to be poorly mineralized, thus very friable, and fusion ohscured articulation in the glyptaled specimens. Bones of the head are generally crushed and those of the hyoid bar, branchial basket, and most of the splanchnocranium and opercular series are either absent or not discernible. All specimens are housed at the South African Museum (SAM). Terminology follows that of standard ichthyology (Hubbs & Lagler, 1958) with one explanation: the dorsal and anal fin origins are said to be associated with a numbered vertebra; this is the centrum whose neural or haemal arch lies immediately anterior to the first fin ray' s pterygiophore ( Anderson, 1988); this feature was distorted in many specimens on fossilization, but restoration was possible for some. All drawings were made with the aid of a camera lucida. Stompooria gen. nov. TYPE SPECIES: Stompooria rogersmithi, sp. nov. DIAGNOSIS: An osmeroid fish (sensu Johnson and Patterson, 1996) with dorsal and anal fins in apposition; first dorsal pterygiophore associated with vertebra 18; anal fin associated with vertebra 19; all neural spines anterior to dorsal fin origin fused to vertebral centra; posttemporal and supracleithrum without connections to seismosensory head canals; neural spine of second and third preural centra morphologically similar; two uroneurals; five hypurals; two epurals; neural arch of first preural/first ural centrum fused with centrum and free of first uroneural; no dorsal outgrowth on first uroneural; principal caudal fin rays 9 + 9; opercle reduced above the hyomandibular articulation; and absence of palatine teeth, maxillary teeth, a mesocoracoid, and supramaxilla. ETYMOLOGY: The name derives from the farm Stompoor where the ancient lake fauna was discovered. Gender feminine. Stompooria rogersmithi sp. nov (Figs. 1-5) Figure 1. Photograph (above) and diagrammatic drawing (below) of holotype of Stompooria rogersmithi, 38.3 mm TL. HOLOTYPE: SAM-K6481a+b (whole counterpiece 6481b, 38.3 mm TL); Stompoor crater core 109, depth 93.98 m; ca. 30 11.5'S, 21 52.2'E; early Maastrichtian age; R.M.H. Smith and A. Crean, Jan. 1984. PARATYPES:(collected with holotype, core depths varying): SAM-K647 5a+b (whole counterpiece K6475a, 43.5 mm TL; half fish K6475b, 28.3 mm), depth 91.12 m. SAM-K6476 (2, both whole, 23.9 and 26.4 mm TL), depth 91.13 m. SAM-K6478 (whole, 34.3 mm TL), depth 92.74 m. SAM-K6487 (whole, 55.8 mm TL), depth unknown. SAM-K6488 (whole, 31.0 mm TL), depth unknown. SAM-K8198 (posterior half, 25.5 mm), depth 42.05 m. SAM-K8246 (whole, 31.5 mm TL, plus partial counterpeice, 18.5 mm), depth 60-66 m. SAM-K8250 (whole, 50.0 mm TL), depth unknown. SAM-K8252 (whole, 49.1 mm TL), depth unknown. SAM-K8253 (whole, 48.3 mm), depth unknown. ADDITIONAL MATERIAL: (collected with types, core depths varying): SAMK6449a+b (2; midbody, 6.8 mm; head + midbody, 19.7 mm), depth 43.11 m. SAM-6457a+b (whole, 25.4 mm TL on counterpiece a; 26.4 mm on counterpiece b), depth 54.42 m. SAM-K6460 (whole, 37.3 mm TL), depth 58.69 m. SAMK6462 (whole, 28.1 mm TL), depth 60.06 m. SAM-K6463 (2; anterior half, 30.3 mm; anterior half, 29.0 mm), depth 62.52 m. SAM-K6468 (whole, 40.6 mm TL), depth 77.65 m. SAM-K6470a+b (anterior half, 52.1 mm on counterpiece a; 45.2 mm on counterpiece b), depth 80.18 m. SAM-K6471 (midbody, 9.1 mm), depth 86.9 m. SAM-K6472a+b (3; midbody, 8.1 mm; midbody, 12.1 mm; midbody 18.4 mm), depth 86.95 m. SAM-K6474 (3; whole, 23.1 mm TL; whole, 26.5 mm TL; anterior half, 25.4 mm), depth 91.1 m. SAM-K6479 (whole, 31.8 mm TL), depth 92.98 m. SAM-K6480 (whole, ca. 33 mm [core cracked]), depth 92.9 m. SAMK6483 (2; whole, 21.5 mm TL; anterior half, 18.1 mm), depth 94.45 m. SAMK6484 (whole, 32.4 mm TL), depth 94.46 m. SAM-K6485 (whole, 48.0 mm TL), depth 94.66 m. SAM-K8199 (whole, 39.0 mm), depth 42.08 m. SAM-K8246 (anterior half, 17.7 mm), depth 60-66 m. SAM-K8247 (2, head region, 9.3 mm; whole, 24.9 mm TL), depth 60-66 m. SAM-K8248a+b (midbody, 36.5 mm), depth unknown. SAM-K8249 (midbody, 25.3 mm), depth unknown. SAM-K8251 (whole, 42.6 mm TL), depth 60-66 m. DIAGNOSIS: As for genus DESCRIPTION: Neurocranium crushed in all material where present, however details of frontals, parasphenoid and top of occipital region allow reconstruction of its basic shape. Parasphenoid wing low, rectangular. Vomer and ethmoid region not readily discernible. Bones of splanchnocranium fragmented, with portions of quadrate and posterior end of articular seen in some specimens. Two elongate infraorbitals coursing posteroventrally present above mesopterygoid (endopterygoid), which has slipped posteroventrally onto the quadrate in SAM-K8253 (Fig. 2); no other infraorbitals are preserved. Mesopterygoid with low medial ridge, suggestive of a bracing structure for tooth row, though no teeth observed. Jaw bones observable in several specimens, with short row of dentary tooth sockets observed. Maxilla without teeth (Fig. 3A). Premaxilla obscured in all material, however, alveolar process not greatly extended under maxilla. Entire palatine present in some specimens, without teeth or tooth sockets (Fig. 3B). Figure 2. Restoration of relationships of mesopterygoid (MES), first and second infraorbitals (INF), quadrate (QUAD) and dentary (DEN) bones in head of Stompooria rogersmithi, SAM-K8253. Figure 3. Isolated bones of Stompooria rogersmithi. A) Left maxilla of SAM-K8251. B) Right palatine of SAM-K8253. C) Right opercle of SAM-K6481b. D) Right preopercle of SAM-K8246. E) Pectoral girdle of SAM-K8246, cleithrum (CL) body restored after SAM-K6487 in part; posttemporal (PT) moved dorsoanteriorly to in vivo position; actinosts (ACT) somewhat disto,~ed and moved posteriorly off cleithrum; SCL: supracleithrum; SCAP: scapula; postcleithrum unknown. Pectoral girdle relatively complete in SAM-K8246 (Fig. 3E), only impression in sediment of coracoid preserved. Mesocoracoid absent. Posttemporal elongate, no ventral (intercalar) ramus. No foramina for passage of postorbital head canal in posttemporal or supracleithrum. Supracleithrum stout, irregular. Cleithrum narrow, with open trough (best observed in holotype). Four rectangular actinosts. Scapula ovoid or triangular, its foramen enclosed by bone. Opercle triangular, dorsal edge canted posteroventrally and below level of hyomandibular articulation (Fig. 3C). Preopercular branch of cephalic lateralis system passing through open groove in preopercle (Fig. 3D). Subopercle and interopercle not preserved. Vertebrae 18 + 23-24 = 41-42 (in three fish). Pleural ribs well developed; epipleurals and epicentrals not evident in present specimens. Epineurals short, at least some evident in most specimens, extending to interneural space between vertebrae 18 and 19. Caudal vertebrae compressing anteroposteriad. All vertebrae equiamphicoelous. Dorsal fin origin associated with vertebra 18. Dorsal fin rays 10-11, usually 11. Anal fin origin associated with vertebra 19. Anal fin rays 10-12, usually 11. In specimens with D 11, A 11, both dorsal and anal fin pterygiophore interdigitation formula 2:2:2:1:1:1:1:1. First element in both dorsal and anal fins short, unbranched, unsegmented; second element unbranched but segmented, remaining rays branched and segmented. Pectoral fin wedge- shaped, rays 11-12, dorsalmost two and ventralmost unbranched. Pelvic fin rays 6-7, reaching posteriorly to anal fin origin; distalmost two and proximalmost rays unbranched. Caudal skeleton with first preural and first ural centre fused, second ural centrum separate. Neural arch of first preural/first ural centre fused to centrum (Fig. 4). Two uroneurals and two epurals. Five hypurals, with parhypural not fused with i~rst hypural. Principal caudal rays 18, dorsal- and ventral-most two unbranched. Procurrent caudal rays 10-12 (upper) and 10-13 (lower). Figure 4. Diagrammatics of two caudal skeletons of Stompooria rogersmithi. A) SAM-K6488b; hypural 3 missing; parhypural, hypural 1 and hypural 5 broken; first uroneural presumed slipped anteriorly to its position in vivo. B ) SAM-K6481 b; epurals and second uroneural missing. Right side drawn in left view. Scales cycloid, radii absent, averaging 0.8 mm in length by 1.0 mm high in SAM-K6487 (55.8 mm TL) under dorsal fin base. Scale outlines and impressions of broadest circuli (mostly at mid-scale regions) visible in best specimens; best preserved scales show anterior margins very slightly scalloped. Blackish pigment spots of various sizes (some quite large) apparently represent chromatophores (see Smith, 1995); these generally not evident in most specimens. ETYMOLOGY: Named in honour of Roger M.H. Smith, South African Museum, collector and interpreter of the crater lake ecosystem.
Fink (1984) restricted the previously constructed Salmoniformes (sensu Greenwood et al., 1966; Rosen, 1974) to one family, Salmonidae. Both Fink (1984) and Rosen (1985) had doubts about the construction of the salmoniform suborder Osmeroidei of the time (which included galaxiids). Begle (1991, 1992) cladistically examined the relationships of two groups he considered close, the argentinoids and osmeroids, and erected the order Osmeriformes (as "Osmerae") to include both groups as suborders. Within Osmeroidei Begle included four "southern smelt" families within the superfamily Galaxioidea. Patterson and Johnson (1995) challenged the value of several of Begle's characters and/or their states and concluded that "there is, as yet, no unequivocal support for argentinoid-osmeroid relationships, and we offer the alternative that salmonoids are the sister group of osmeroids..." Johnson and Patterson (1996) critically analyzed Begle' s work and found it to be unreliable, as characters had been misinterpreted or miscoded, and they reclassified the osmeroids with the salmonoids as order Salmoniformes. Derived characters produced two cladograms, one optimization favoring forward changes and another favoring reversals. This resulted in homoplastic characters partially defining different clades between the two methods (Johnson and Patterson, 1996, Fig. 19). Those derived characters that are verifiable in Stompooria from their optimization favoring forward change for Osmeroidei are (numbers in parentheses are their character numbers): 1) opercle below level of hyomandibular articulation (37) (homoplasy with Galaxioidea in the other analysis); 2) first ural and first preural centra fused (65); 3) two epurals (68) (homoplasy with Galaxiidae in the other analysis); 4) no postcleithrum (80). Derived characters verifiable in Stompooria for Galaxioidea in Johnson and Patterson's optimization favoring forward change are: 1) maxilla without teeth (29); 2) supramaxilla absent (30); [the following 5 characters are homoplasies with Salangidae in the other analysis]: 3) two uroneurals (72); 4) posttemporal without ossified ventral (intercalar) ramus ( 7 8 ); 5 ) mesocoracoid absent ( 81 ); posttemporal without connection to seismosensory system (99); 6) supracleithrnm without connection to seismosensory system (100). Derived characters verifiable in Stompooria for Johnson and Patterson's expanded Galaxiidae in the analysis favoring forward change are: 1) mesopterygoid (endopterygoid) teeth in single row (presumed; see Description) (23) (homoplasy with most northern smelts in the other analysis); 2) all neural arches anterior to dorsal fin fused to centra ( 55 ); 3 ) neural spine of second preural vertebra similar in form to neural spine of third preural vertebra (63); 4) five hypurals (67); 5) no dorsal outgrowth on first uroneural (70); 6) no palatine teeth (not coded, appendix 1; C. Patterson, in litt.). In addition, Stompooria possesses epineurals, interpreted by Johnson and Patterson as a reversal partially defining their tribe Galaxiini (traditional Galaxiidae). Insufficient observation of characters, many of them based on soft tissues, makes the assignment of Stompooria to a family group as defined by Johnson and Patterson (1996) difficult. Stompooria is excluded from Retropinnidae in having the parhypural and first hypural unfused (character 66), as well as characters noted by McDowall (1969) separating galaxiids and retropinnids such as five hypurals, dorsal fin over anal fin, and (apparent) absence of palatine teeth, although Stompooria has 9+9 principal caudal rays, as in retropinnids. As noted above, Stompooria seems to fit within Johnson and Patterson' s expanded Galaxiidae (includes aplochitonids and the monotypic Lepidogalaxias salamandroides), albeit at a primitive level, a reasonable conclusion given its age. Those character states that indicate its more plesiomorphic status include: unmodified pectoral radials, presence of scales, normal neural spine of the first preural vertebra, two epurals, relatively few vertebrae, and the dorsal and anal fins originating comparatively forward, apparently as a result of its few precaudal vertebrae. Stompooria has 41-42 vertebrae while most extant galaxiids have vertebral counts in the mid 50s to mid 60s (Allen, 1989; McDowall, 1990). Counts for Galaxiella (36-43) and Paragalaxias (37-44) may represent atavisms (loss of vertebrae) in these genera, as I have reported on in other fishes (Anderson, 1994). The far forward dorsal fin position in the extant Paragalaxias is reminiscent of that in Stompooria, although the anal fin is posterior, thus the dorsal/anal relationship is more like that of retropinnids. The unpaired fin/vertebral condition in extant galaxiids may have been derived from the condition observed in Stompooria by a concomitant increase in precaudal vertebrae with a decrease in caudal vertebrae, which I see as a particular adaptation to reduce drag in dart-swimming in these fishes (Alexander, 1967). The absence of scales is generally considered a derived condition in fishes. The plesiomorphic retention of scales in Stompooria is not surprising, but their presence in Lepidogalaxias in Johnson and Patterson' s phylogeny requires scale loss a minimum of twice (once for tribes Aplochitonini and Galaxiini and once for Lovettia), or that scale loss occurred once in these fishes and Lepidogalaxias has atavistically re-acquired them. The latter possibility seems unlikely to me. Finally, it should be noted that the apparent absence of epipleural bones (Johnson and Patterson's tribe Galaxiini; character 59) in Stompooria may be a preservation artifact. The relationship of Stompooria to other known salmoniform fossils from the Late Cretaceous (Patterson, 1970) is remote. As for comparison with other galaxioid fossils, McDowall (1976) and McDowall and Pole (1997) reported Galaxias from New Zealand' s Miocene that are essentially comparable to extant forms. I thank R.M.H. Smith, A. Crean and S. Kaal, South African Museum, Cape Town for curatorial assistance and advice. Mrs. E. Heemstra, J.L.B. Smith Institute, completed my original sketches. Early drafts of the manuscript were improved with the kind assistance of J.A. Long, R.M. McDowall, C. Patterson and P.H. Skelton. ALEXANDER, R.M. 1967. Functional Design in Fishes. Hutchinson University Library, London. 160 pp. ALLEN, G.R. 1989. Freshwater fishes of Australia. T.F.H. Publications, Neptune City, USA. 240 pp. ANDERSON, M.E. 1988. Studies on the Zoarcidae (Teleostei: Perciformes) of the southern hemisphere. I. The Antarctic and subantarctic. Biol. Antarctic Seas, XIX, Antarct. Res. Ser. 47: 59-113. -- 1994. Systematics and osteology of the Zoarcidae (Teleostei: Perciformes ). Ichthyol. Bull. J.L.B. Smith Inst. 60: 1-120. BEGLE, D.P. 1991. Relationships of the osmeroid fishes and the use of reductive characters in phylogenetic analysis. Syst. Zool. 40(1): 33-53. -- 1992. Monophyly and relationships of the argentinoid fishes. Copeia 1992(2): 350-366. FINK, W.L. 1984. Basal euteleosts: relationships. In: Moser, H.G. et al. Ontogeny and Systematics of Fishes. Amer. Soc. Ichthyol. Herpetol. Spec. Publ. 1: 202-206. GREENWOOD, P.H., D.E. ROSEN, S.H. WEITZMAN & G.S. MYERS. 1966. Phyletic studies of teleostean fishes, with a provisional classification of living forms. Bull. Amer. Mus. Nat. Hist. 131(4): 339-455. HUBBS, C.L. & K.F. LAGLER. 1958. Fishes of the Great Lakes region. Cranbrook Institute of Science, Bulletin 26, Bloomfield Hills, xiii+213 pp. JOHNSON, G.D. & C. PAT'I'ERSON. 1996. Relationships of lower euteleostean fishes. In: Stiassny, M.L.J., L.R. Parenti & G.D. Johnson (eds.), Interrelationships of Fishes. Academic Press, London, xiii + 496 pp. McDOWALL, R.M. 1976. Notes on some Galaxias fossils from the Pliocene of New Zealand. J. RoyaI Soc. New Zealand 6(1): 17-22. -- 1990. New Zealand freshwater fishes: A natural history and guide. 2nd ed. Heinemann Reed, Auckland. 553 pp. McDOWALL, R.M. & M. POLE. 1997. A large galaxiid fossil (Teleostei) from the Miocene of central Otago, New Zealand. J. Royal Soc. New Zealand 27(2): 193-198. PATTERSON, C. 1970. Two Upper Cretaceous salmoniform fishes from the Lebanon. Bull. British Mus. (Nat. Hist.), Geol. 19(5): 207-296, pls. 1-5. PATTERSON, C. & G.D. JOHNSON. 1995. The intermuscular bones and ligaments of teleostean fishes. Smithson. Contr. Zool. (559): 1-83. ROSEN, D.E. 1974. Phylogeny and zoogeography of salmoniform fishes and relationships of Lepidogalaxias salamandroides. Bull. Amer. Mus. Nat. Hist. 153(2): 265-325. ROSEN, D.E. 1985. An essay on euteleostean classification. Amer. Mus. Novit. (2827): 1-57. SMITH, R.M.H. 1986. Crater lakes in the age of dinosaurs. Sagittarius 1(1): 10-15. -- 1988. Palaeoenvironmental reconstruction of a Cretaceous crater-lake deposit in Bushmanland, South Africa. In: Heine, K. (ed.), Palaeoecology of Africa and the Surrounding Islands 19:, 27-41. -- 1995. Life in a prehistoric crater-lake. The Phoenix 8 (2 & 3): 4-6. VAN DIJK, D.E. 1985. An addition to the fossil Anura of southern Africa. So. Afr. J. Sci. 81: 205-206. Copyright 1998 J.L.B. Smith Institute of Ichthyology The following images related to this document are available:Photo images[fs98001a.jpg]Line drawing images[fs98001e.gif] [fs98001d.gif] [fs98001b.gif] [fs98001c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}