 |
search
for |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Special Publication No.62, November 1998
A NEW LUMINOUS CARDINALFISH OF THE GENUS APOGON (PERCIFORMES, APOGONIDAE) FROM THE WESTERN PACIFIC OCEAN
Code Number:FS98003
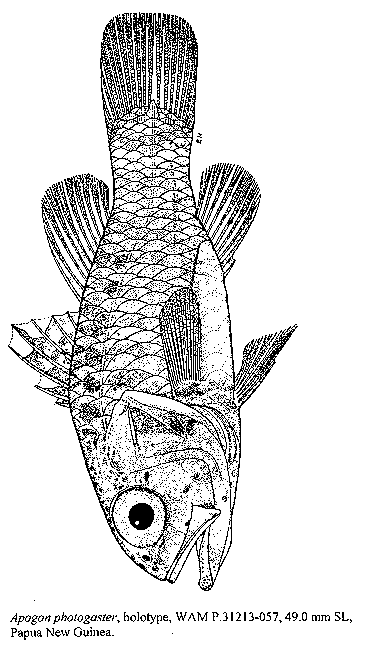
ABSTRACT A new species of cardinalfish, Apogon photogaster is described based on 11 specimens from Madang Lagoon in Papua New Guinea and Galoc Island in the Philippines. This new species has 15 pectoral rays, 2 predorsal scales, 15-18 gill-rakers (8-9 developed), and 7 gill-rakers on the ceratobranchial of the first gill-arch. It has a unique bioluminescent system, including a light organ branching forward from the second loop of the intestine and a diffuser organ along the ventrolateral part of the body. INTRODUCTION Bioluminescence in the Apogonidae was discovered some 50 years ago (Kato, 1947) with the description of the light organs of Apogon truncatus Bleeker, 1854. Eleven years later Iwai & Asano (1958) found the same organs in A. smithi (Kotthaus, 1970). Iwai (1958) was the first to associate the ventrolateral, striated silvery stripe of Siphamia with bioluminescence. Further studies have demonstrated the existence of two different bioluminescent systems in the Apogonidae. Luminous species of Apogon have a chemical system in which light is produced by a luciferin-luciferase reaction (Haneda et al., 1958). In Siphamia the light is emitted by luminous bacteria found in the light organ of these fishes (Haneda, 1965; Iwai, 1971). Recently, Gon (1996) included the luminous species of Apogon in subgenus Jaydia (Smith, 1961). In this paper we describe a new luminous species of Apogon, subgenus Jaydia, from specimens collected by G.R. Allen in Papua New Guinea and the Philippines. METHODS Type specimens were lodged with the following institutions: Bernice P. Bishop Museum, Honolulu (BPBM); Muséum National d'Histoire Naturelle, Paris (MNHN); J.L.B. Smith Institute of Ichthyology, Grahamstown (RUSI); Western Australian Museum, Perth (WAM); United States National Museum of Natural History, Washington D.C. (USNM). The method of measuring fish follows Gon (1993). Measurements were taken to the nearest .05 mm. Standard length (SL) was measured from the tip of the upper jaw to the end of the hypurals. Counts of pectoral-fin rays and lateral-line scales were made on both sides of the fish. The former count includes the uppermost rudimentary ray. The lateral-line scale count is the number of tubular scales on the body + those extending onto the caudal-fin base. A developed gill-raker is higher than the width of its base. The raker at the angle of the gill-arch is included in the lower-limb count. When a raker is found on the ceratobranchial-hypobranchial joint it is included in the ceratobranchial gill-raker count. Apogon (Jaydia) photogaster, sp. nov. Figure 1. Apogon photogaster, holotype, WAM P.31213-057, 49.0 mm SL, Papua New Guinea. Holotype: WAM P.31213-057, 49.0 mm SL, male, Papua New Guinea, Madang Lagoon, Padoz Reef (510'S, 14549'E); 18-23 m, coral, sand and rubble; rotenone, G.R. Allen & A. Jenkins, 21 October 1996. Paratypes: BPBM 38301, 42.0 mm SL, female, collected with holotype; MNHN 1998-391, 34.1 mm SL, male, collected with holotype; RUSI 57596, 41.4 mm SL, male, collected with holotype; RUSI 57597, 47.4 mm SL, collected with WAM P.30358-001 (cleared & stained); USNM 348213, 43.6 mm SL, female, collected with holotype; WAM P.30358-001, 29.8 mm SL, female, same locality as holotype, 26-30 m, small coral head and silt bottom, rotenone, G.R. Allen, 28 October 1991; WAM P.31213-001, 3: 31.4-48.8 mm SL, 2 females 1 male, collected with holotype; WAM P.31394-001, 2: 65.5-72.4 mm SL, females, Philippines, Calamian Group, SW end of Galoc Island, 1156.81'N, 11949.47'E, 18 m, rotenone, G.R. Allen, 12 March 1998. DIAGNOSIS: Dorsal rays VII + I,9; anal rays II,8; pectoral rays 15; lateral-line scales 24 + 3 on caudal fin; predorsal scales 2; total gill-rakers on first gill-arch 4 + 11-14; developed gill-rakers 1 + 7-8; gill-rakers on ceratobranchial 7-8 (usually 7). Bioluminescent system including a light organ branching forward from second loop of intestine and a diffuser organ. Body whitish to pale brown with 7-10 faint dark stripes; anterior end of branchial basket and base of tongue blackish; peritoneum and part of intestine beyond the light organ with dark spots. DESCRIPTION: Data in parentheses refer to paratypes (when different from the holotype), 11 measured and counted. Proportional measurements of the holotype and selected paratypes are shown in Table 1. Pectoral rays 15; lateral-line scales 24 to end of hypural plate plus 3 beyond it; predorsal scales 2; total gill-rakers on first gill-arch 4 + 13 (11-14); developed gill-rakers 1 + 7 (7-8), 3 of 11 paratypes with 7 rakers on lower limb; developed gill-rakers on ceratobranchial 7 (7-8), 1 paratype with 8 rakers. Body oblong and compressed, its depth 2.7 (2.7-2.9) in SL and its width 1.8 (1.7-2.2) in the depth; head length 2.3 (2.2-2.6) in SL; snout length 4.8 (4.3-5.8), eye diameter 3.2 (2.5-3.3), interorbital space 5.3 (4.85-6.1), upper-jaw length 2.2 (1.9-2.2), lower-jaw length 1.8 (1.7-1.95), all in head length. Preopercle edge with small serrae on ventral part, around angle, and lower half of vertical part; preopercle ridge smooth; posttemporal with four minute serrae, but smooth in 9 of 11 paratypes. Posterior nostril small (3-4 times in pupil) and oblong, placed at a distance equal to its length in front of eye and at level of middle of eye; anterior nostril smaller, round, with a short tube and situated about mid-way between rear nostril and upper lip, or slightly closer to the latter. First infraorbital pore slightly smaller than rear nostril; second and third infraorbital pores, located on ventral edge of infraorbital bone, close together and larger than first pore; first supraorbital pore larger than rear nostril, just above upper lip, slightly slanting up toward anterior nostril; second supraorbital pore as large as nostril, medial to the latter; third supraorbital pore just in front of a vertical through middle of eye and only slightly larger than small secondary pores near it. Mouth oblique, large, the maxilla reaching to below posterior edge of pupil; maxilla depth 3.3 (3.2-3.8) in length of upper jaw, its posterior edge indented; both jaws with a polyserial band of small, conical teeth; upper jaw band slightly tapered at both ends; lower jaw band widest at symphysis, gradually tapering to 1-3 series posteriorly; vomer with 1-2 series of minute teeth; palatines toothless, but some paratypes have a few minute teeth on anterior part of these bones. Dorsal profile of head slightly convex; origin of first dorsal, pectoral, and pelvic fins more-or-less on the same vertical; first dorsal spine 2.3 (2.0-2.8) in second spine; second dorsal spine 5.3 (4.3-5.8), third dorsal spine 2.7 (2.2-2.9), fourth dorsal spine slightly longer 2.5 (2.2-2.7), spine of second dorsal fin 3.2 (2.8-3.5), longest dorsal ray (second or third) 1.9 (1.5-2.1), all in head length (in 3 paratypes the third dorsal spine is slightly longer than the fourth, and in 3 other fish these spines are subequal); anal-fin origin at level of base of fourth ray of second dorsal fin; first anal spine 3.7 (2.6-3.8) in second spine; second anal spine 3.9 (3.2-4.2) and longest anal ray (second or third) 2.0 (1.7-2.3) in head length; pectoral length 4.3 (3.9-4.6) and pelvic length 3.8 (3.4-4.1) in SL; pelvic spine 3.0 (2.35-3.1) in head length and 1.9 (1.55-2.1) in its fin length; distance from snout to first dorsal-fin origin 2.3 (2.2-2.3), to second dorsal-fin origin 1.6 (1.5-1.6), to pelvic insertion 2.45 (2.4-2.8), and to anal-fin origin 1.5 (1.4-1.6), all in SL; caudal peduncle depth 1.15 (1.1-1.6) in its length, the length 5.4 (4.5-5.6) in SL; distance from anus to anal-fin origin 6.5 (5.25-10.8) in distance from pelvic insertion to anal-fin origin, and the latter distance 3.5 (2.9-3.6) in SL. The light organ is an anterior cylindrical protrusion branching from the second loop of the intestine and fitting into an extension of the body cavity above the pelvic girdle (Fig. 2); the diffuser organ is made of translucent muscle and appears externally as a fairly wide ventrolateral silvery stripe ending at about middle of anal-fin base. Figure 2. Ventral view of the intestine of Apogon photogaster, showing the light organ ( pale cylindrical extension in front of spotted gut) pulled out of the anterior extension of the body cavity (black oval) into which it fits. Colour in alcohol, pale brown on body, darker on head; opercle, cheek and lateral parts of snout peppered with small melanophores; melanophores are more widely spread on top of snout and head, some are stellate; several irregular dark brown spots on cheek behind and below eye; in the smallest paratype (29.8 mm SL) these spots are part of cheek and temporal stripes radiating from the eye; gill-arches, gill-rakers and vomer with small blackish spots; anterior end of branchial basket and base of tongue blackish; body with 8 faint bars (7-10 in paratypes); dorsal ends of first 4 bars darker, appearing as a series of blackish spots above lateral line; a wide silvery band along ventral part of body from pectoral and pelvic bases to middle of anal-fin base; leading edge and distal part of first dorsal fin dark brown; second dorsal fin pale, except for dusky spots between spine and anterior 2-3 rays; leading edge of pelvic fins dark brown; anal fin brown anteriorly, the last five rays dusky only on distal part; caudal fin dusky; peritoneum with fairly dense black spots of various sizes; part of intestine behind light organ with blackish spots. Colour in life (from colour slide by G.R. Allen) similar, but with pinkish hue. REMARKS: The bars on the body are more distinct in smaller fish. The amount of dark pigment on the fins is variable, but seems to increase with size. The anterior end of the branchial basket and base of tongue are blackish in all fish regardless of size. In the largest two specimens (65 & 72 mm, from the Philippines) the fins (except pectorals) are darker. The bioluminescent system of photogaster is unique because it combines components found in two different apogonid systems. The intestinal light organ is typical of the chemical light system of subgenus Jaydia, and the ventrolateral diffuser organ is typical of the bacterial light system of genus Siphamia. These two features distinguish photogaster from all known apogonid species. Certain members of Jaydia have an intestinal light organ, but not the diffuser organ. Unlike the diffuser organ of photogaster, this organ in Siphamia is striated and extends onto the caudal peduncle, gradually tapering posteriorly. In addition, the diffuser organ of photogaster lacks a layer of melanophores external to the epimysium of the organ of Siphamia. Apogon photogaster belongs in the lineatus group of Jaydia. This species group also includes striatus, striatodes, lineatus and novaeguinae (Gon, 1996). Within this group photogaster is more closely related to striatus and striatodes. All three species have 15 pectoral rays, an intestinal light organ, and a dark spotted peritoneum. In lineatus and novaeguinae the peritoneum is pale (sometimes with a few scattered dark spots) and both species have no light organ. A. photogaster has fewer developed gill-rakers (8-9) than the other four species of the lineatus group (10-15 rakers). It also resembles smithi, hungi and truncatus of the truncatus group (Gon, 1996), but these species have 16-17 pectoral rays, 3-5 predorsal scales, and 8-9 gill-rakers on the first ceratobranchial (in photogaster these characters are 15, 2, and 7, respectively). The specimens from Madang, Papua New Guinea, were collected from a patch reef in a sheltered lagoon environment at depths of 18-30 m. The patch reef forms part of a complex of several similar reefs, which rise from the lagoon floor in 35-40 m depth and approach to within 2-3 m of the surface. The reefs are roughly circular with their shallow tops occupying up to 2-3 acres. Specimens were taken on a relatively steep slope near the base of the reef in an area of heavy sedimentation characterized by occasional outcrops of live and dead coral. The fish are apparently cryptic or nocturnal as none were sighted until rotenone was dispersed. This species is named photogaster (Greek for luminous belly) in reference to the ventral bioluminescent system. Table 1. Proportional measurements of the holotype and six (of 11) paratypes of Apogon photogaster expressed as a percentage of standard length ( = damaged)
We thank J.T. Eastman of Ohio University, U.S.A., for histological confirmation of the diffuser organ, and E. Heemstra of the J.L.B. Smith Institute of Ichthyology for the artwork presented in this paper. LITERATURE CITED BLEEKER, P. 1854. Specierum piscium javanesium novarum vel minus diagnoses adumbratae. Nat. Tijdschr. Neder.-Indië 7: 415-448. GON, O. 1993. Revision of the cardinalfish genus Cheilodipterus (Perciformes, Apogonidae), with description of five new species. Indo-Pac. Fishes (22): 1-59. GON, O. 1996. Revision of the cardinalfish subgenus Jaydia (Apogonidae, Apogon). Trans. Roy. Soc. S. Afr. 51: 147-194. HANEDA, Y. 1965. Observation on a luminous apogonid fish, Siphamia versicolor, and on others of the same genus. Sci. Rep. Yokosuka City. Mus. (11): 1-12. HANEDA, Y., F.H. JOHNSON & E.H.C. SIE. 1958. Luciferin and luciferase extracts of a fish, Apogon marginatus, and their luminescent cross reaction with those of a crustacean, Cypridina hilgendorfi. Biol. Bull. 115: 236. IWAI, T. 1958. A study of the luminous organ of the apogonid fish Siphamia versicolor (Smith and Radcliffe). J. Wash. Acad. Sci. 48(8): 267-270. IWAI, T. 1971. Structure of luminescent organ of apogonid fish, Siphamia versicolor. Jap. J. Ichthyol. 18(3): 125-127. IWAI, T. & H. ASANO. 1958. On the luminous cardinal fish, Apogon ellioti Day. Sci. Rep. Yokosuka City. Mus. (3): 5-13. KATO, K. 1947. A new type of luminous organ of fish. Zool. Mag. 57(12): 195-198. (In Japanese). KOTTHAUS, A. 1970. Fische des Indischen Ozeans, Ergebnisse der ichthyologischen Untersuchungen während der Expedition des Forschungsschiffes "Meteor" in den Indischen Ozean, Oktober 1954 bis Mai 1965. A. Systemtaticher Teil, VIII, Percomorphi (2). "Meteor" Forsch. Ergebnisse Series D, 6: 56-75. SMITH, J.L.B. 1961. Fishes of the family Apogonidae of the Western Indian Ocean and the Red Sea. Ichthyol. Bull. Rhodes Univ. (22): 373-418. Copyright 1998 J.L.B. Smith Institute of Ichthyology The following images related to this document are available:Line drawing images[fs98003b.gif] [fs98003a.gif] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}