 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Journal of Food Technology in Africa, Vol. 6, No. 1, Jan-Mar, 2001, 13-17 Genotypic variation of Kenyan tomato (Lycopersicon esculentum L.) germplasm
S. G. Agong*, S. Schittenhelm**, W. Friedt*** *Department of Horticulture, JKUAT, P. O Box 62000 Nairobi, Kenya Code Number: ft01005
Abstract
Genotypic variation of Kenyan tomato (Lycopersicon esculentum L.) germplasm Systematic genotypic analysis of Kenyan tomato germplasm was carried out in order to delineate potential variability based on various morphological, agronomic and biochemical traits. Both landraces and market cultivars were examined with a view to facilitating tomato improvement. In an experiment conducted in 1993 in a glasshouse at the Federal Agriculture Research Centre (FAL), Germany, 26 tomato landraces and nine market cultivars were investigated using block design. Analysis of variance clearly illustrated a large variation for all the quantitative traits. Landraces on average produced more fruit per plant (90) but of a smaller size than the market cultivars (19). However, market cultivars had a superior average fresh fruit weight of 56.5g while the landraces registered on average 40.6 g. Multiple correlation analysis confirmed the superiority of landraces for trait of fruit quality and a strong negative association between fruit weight. Limited structure groupings were detected on the basis of a principal components analysis. Using this method, processing and fresh tomato cultivars within the germplasm could be clearly separated on the basis of fruit characters. Furthermore, this analysis distinguished a few landraces from the market cultivars, although closer phylogeny through introgression was highly suspected. Within the landraces, the yellow-cherry types were distinct from all the others, On the basis of this study, the use of more prolific landraces, in terms of number of fruit as well as actual fruit yield, would be desirable for intensive and continuous production of tomatoes. Key Words: Genetic diversity, landraces, Lycopersicon esculentum, phylogenic relationships, principal components analysis, Tomato Introduction
With the increasing need of consumers for both quality and diversity of tomato products, there is a need to extensively collect, exploit and evaluate unknown tomato germplasm. Tomato continues to play a key horticultural role in Kenya and its improvement would enhance agricultural productivity, alleviate poverty and facilitate food security (Agong and Schittenhelm 1993; Agong et al. 1997). However, most of the tomato germplasm in the country is largely undocumented and has unknown morphological, agronomic and biochemical attributes. Tomato is continuously introduced and grown in all ecological zones where arable agriculture is practicable. This tendency has fuelled the extensive cultivation of various tomato cultivars with unclear documentation (Agong and Schittenhelm 1993).
Systematic study and characterisation of tomato germplasm is of great importance for current and future agronomic and genetic improvement of the crop. Furthermore, if an improvement programme is to be carried out evaluation is imperative, in order to understand the genetic background and the breeding value of the available tomatoes.
Morphological, agronomic as well as biochemical parameters have been widely used in the evaluation of various crops (Rick and Holle 1990; Weber and Wricke 1994; Kaemmer et al. 1995). Exploitation of such traits increases our knowledge of the genetic variability available and strongly facilitates breeding for wider geographic adaptability, with respect to biotic and abiotic stresses. In addition, genetic diversity needs to be described and measured if it is to be effectively incorporated into breeding strategies and management of plant genetic resources.
The objective of this study, therefore, was to examine the variation in tomato germplasm based on the morphological, agronomic and biochemical traits in the landraces, as well as in market cultivars, with an ultimate view of identifying potential accessions to improve tomato production. This study also aimed to generate data to increase understanding of the phylogeny of the Kenyan tomato germplasm to improve effective management. Materials and Methods
Tomato (Lycopersicon esculentum L.) accessions collected from different parts of Kenya, as described by Agong and Schittenhelm (1993) and Agong et al. (1997), were used in this study. Germplasm was comprised mostly of landraces grown by mainly small-scale farmers over several years, and collected mainly in the western, central, eastern and coastal regions of Kenya, These areas differ greatly in their agro-ecological and ethnic compositions.
Morphological, agronomic and biochemical characterization of the tomato germplasm was conducted with the hypothesis that any differences among the tomato accessions would be due to the genetic differentiation therein and not solely to phonotypic plasticity, given the diverse environmental differences between the collection sites (Agong and Schittenhelm 1993). Using a four-replicate randomized complete block design, a pot experimental study under a controlled glasshouse environment was conducted from February to August 1993 at FAL. For each replicate, 12 plants per accession were studied, in the experiment 35 accessions were used, including three of German origin used as control. The tomatoes were grown in an organically enriched Volkenrode compost soil following standard horticultural practice, as described in Agong et al. (1997). Throughout the experiment, glasshouse temperatures were kept at 15°C at night and 25°C during the day and relative humidity maintained at 70%. The seedlings were pricked out into 26 cm plastic pots four weeks after emergence and cultured up to bright-red fruit maturity. The parameters scored during vegetative growth and through fruit maturity included: total fruit weight per plant of mature fruits (g FFW), dry fruit weight per plant of mature fruits (g DFW), plant height at fruit maturity (cm PH), mature fruit dry matter content (% DC), dry fruit weight of mature fruit (g SW), electrical conductivity (dS/m EC), pH value (pH), Brix (5 BRI), titratable acidity (% TA), citric acid (mM/LCA), malic acid (mM/L MA), fructose (% FRU) and glucose (% GL) of mature fruit. Fruit juice extracts from each tomato accession were obtained and stored at -20°C for the chemical analysis. From each of the 12 plants per accession in every replication, 5g of juice extract was obtained to provide a 60 mixed juice sample for use in the biochemical analyses.
Sampling was done carefully to ensure that fruit from all the accessions was at an approximately similar physiological maturity (bright red ripe). Thawed juice samples were vigorously shaken and used to determine the electrical conductivity and pH values according to the procedures of Agong et al. (1997). The percentages of total soluble solids was measured by the use of sucrose hand refractometer (model HRN-20 of Kruss-W.S.R. Tokyo).
Sugars (glucose and fructose) and organic acids (citric and malic) were analyzed from the homogenously mixed fruit-juice extract using high performance liquid chromatography (HPLC) as described by Mitchell et al. (1991). However, the sugars were extracted in distilled water and not in 60% ethanol. The separation of sugars and organic acids was accomplished on an AminexTM HPX-87H 300x 7.8 mm column (BIO-RAD, Richmond, California) with a degassed 0.05 N H2SO4 as the mobile phase (eluant) at a flow rate of 0.4 ml/min. Sugars were detected with a differential refractometer (RI-Ditektor, Knauer) and organic acids by UV absorbance at 210 nm. All samples were run at a constant temperature of 30°C. Titratable acidity was determined using the methods of the National Canners Association (1968) and Agong et al. (1997).
By using the computer program SAS (SAS Institute Inc. 1990) an analysis of variance (ANOVA) was carried out to determine the significance of difference. Multiple correlation and principal components analysis (PCA) were carried out as described by Broschat (1997) on the standardized and normalized mean value of the metric characters and correlation matrices. Results
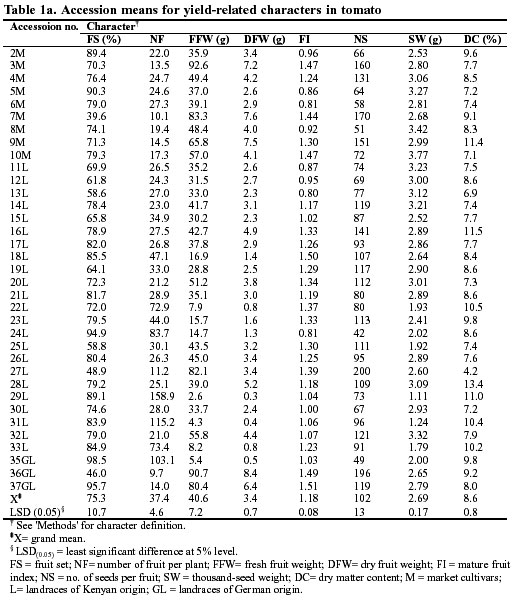
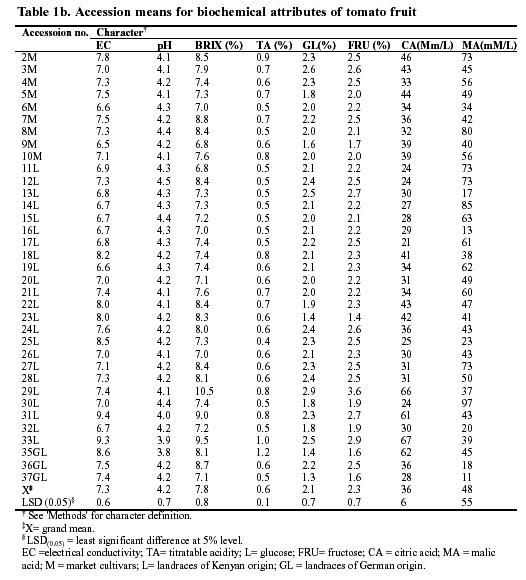
The evaluation of the Kenyan tomato germplasm showed a large and significant variation in the quantitative traits between the accessions (Table 1a, b). For example, the percentage of fruit set was scored at 39.6 and 94.9% for accessions 7 and 24, respectively. The average fresh and dry fruit weight varied notably among the accessions. Most of the landraces gave lower fresh and dry fruit yields than the market cultivars. On the other hand, the landraces displayed superiority with respect to biochemical parameters. For example, landraces 29, 31 and 33 had very high levels of Brix (Table 1b). The electrical conductivity for all accessions ranges between 6.5 and 9.4 dS/m. Similarly, the fruit juice extract of the landraces, particularly accessions 33 and 31, had the highest electrical conductivity.
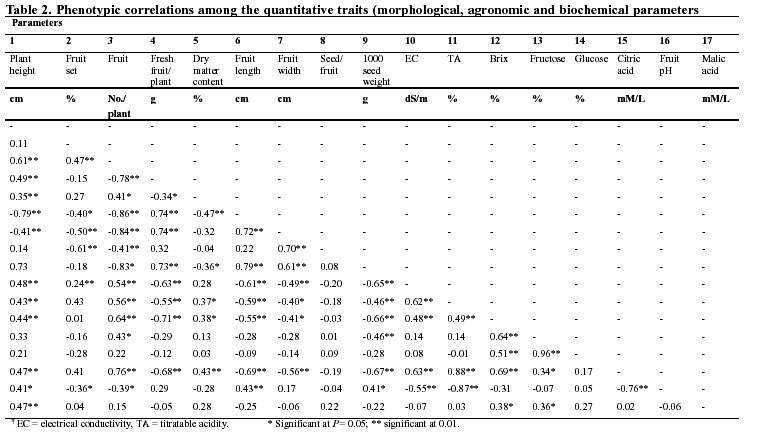
Correlation analysis revealed strong relationships among the biochemical traits (Table 2). As expected, the weight of fresh fruit was negatively associated to the fruit's biochemical contents. Fresh fruit weight also correlated negatively to fruit number per plant and dry matter content. Positive correlationship, however, was observed between fresh fruit weight and fruit width and fruit equatorial length, whereas the number of fruit per plant was negatively correlated for fruit width.
Visual appraisal of the germplasm during the vegetative and the reproductive phases also showed that the accessions were fairly variable. For example, observation of the reproductive parts revealed that accession 33, a red-cherry tomato, had a pin flower form whereas the yellow-cherry and the market types displayed the thrum flower structure (Fig. 1) Similarly the fruit forms differed.
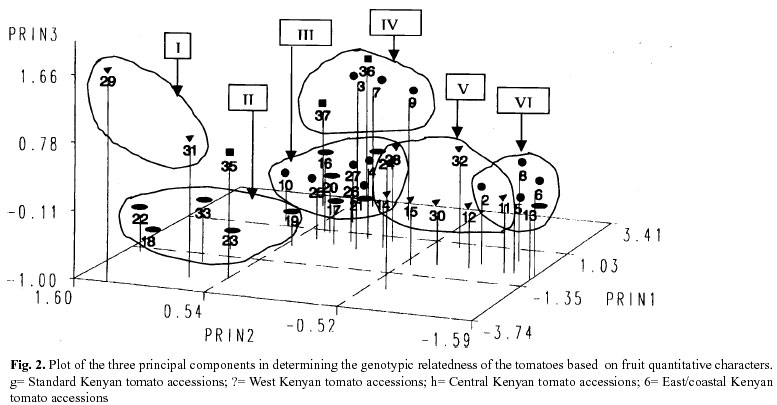
The PCA on 15 unrelated but linearly correlated quantitative characters indicated that the first three principal components were adequate in explaining more than 70% of the phenotypic variation in the tomato germplasm (fig. 2). However, no clear classes could be adduced from the analysis. To some extent it was possible to distinguish between accessions collected in the western, central and coastal areas of Kenya. An approximate classification is (a) landraces with yellow fruit from the coast, (b) landraces with red-cherry fruit from western Kenya, (c) a mixture of coastal, central and western accessions, (d) central accessions with large fruit, (e) coastal accessions with large fruit and (f) a mixture of all accessions. Accessions 35, 36 and 37 were used for control purposes and do not affect the possible genotypic classifications. Market cultivars can be separated into three categories: 4 and 10; 3, 7 and 9; and 2, 6 and 8. Tomatoes for processing (2, 5, 6 and 8) could clearly be separated on the basis of PCA analysis for the fresh market cultivars (3, 4, 7, 9, and 10). Discussion
The heterogeneity observed in the germplasm is largely attributable to the genotypic variability within and between the individual tomato groups. The variation adduced in this study conforms with earlier work on the reaction of this germplasm to salinity stress (Agong et al. 1997). The availability of a base variable population, for example red and yellow cherry fruiting tomato types, is crucial for any significant progress in crop genetic advancement. Genetic improvements of tomato should not only depend on the introduction but also on the gradual development of more closely adapted accessions suited to local conditions (Agong 1995).
On the basis of morphological, agronomic and biochemical data generated in this study on yield and yield-related traits, it is suggested that fruit number per plant and fruit index (length/width), which are closely associated with fresh fruit yield (Table 2), can be used to create a better understanding of diversity in the tomato for yield and crop improvement (Cavicchi and Silvetti 1976). The percentage of fruit set and fruit number per plant were strongly correlated, hence this relationship can be useful for effective pruning management as well as for predicting new selection procedures for crop improvement (Cavicchi and Silvetti 1976).
The pH, total titratable acidity and Brix are vital attributes with respect to the organoleptic quality of tomato fruit (Tigchelaar 1986; Mitchell et al. 1991; Agong 1995). On average, most landraces had a high number of biochemical attributes (Table 1b) and these characteristics are definitely useful where production tends to be for processing. Thus landraces may be a valuable source of germplasm for the improvement of processing tomatoes. The greater commercialization of these landraces would strongly motivate and economically empower small-scale farmers who possess a large portion of the germplasm.
The lack of definitive classification based on the PCA strongly suggested closer phylogenic relationship amongst the tomato accessions (Fig. 2). Fruit characters alone are unlikely to be suitable for evaluating tomato germplasm. The inclusion of more morphological, agronomic and biochemical traits would be appropriate, especially in a multi-trait selection programme for the improvement of horticultural characteristics (Broschat 1979). However, visual appraisal of germplasm during the vegetative and the reproductive phases confirmed genotypic variability within the germplasm (Fig. 1).
These visual features were extremely helpful in genotypic differentiation of the landraces, such as the expression of stigma above the anther come in accession 33, suggesting the close phylogenic relationship of this landrace to the primitive progenitor of tomato (L. esculentin var cerasiformie), known for its out-breeding tendency (Rick 1976; Alcazar-Esquinas 1981). From an evolutionary standpoint, if farmers have been practising methods which encourage intense inbreeding, it is very likely that rare genes will ultimately be expressed, thus exposing the wild phenotypes as observed in accession 33. Modern tomato cultivars comprise strongly self-pollinating types that have a limited chance of cross-pollination.
As expected, correlation analyses revealed that fresh fruit yield was negatively associated to fruit number (Table 2). Thus, if small-scale farmers have been selecting for higher fruit number they might have done so at the expense of improving yields. Most tomato landraces had a higher number of fruit per plant than the market cultivars, confirming their mostly inferior fresh fruit yield in comparison to the market cultivars, as evidenced elsewhere (Agong 1995). Over a long time period the high production of fruit in landraces can substantially benefit urban and peri-urban communities. Thus landraces can effectively be utilized under intensive and continuous tomato production systems.
Most of the biochemical characters were negatively correlated to fresh fruit yield (Table 1b). Therefore, a breeding programme would sacrifice the larger fruit to obtain better quality, particularly when the main objective is to improve the processing quality (Agong et al. 1997). Electrical conductivity, Brix [%], pH value and total titratable acidity are used as criteria to judge the organoleptic and processing qualities of tomato and, therefore, require inclusion into breeding programmes (Mitchel et al. 1991; Agong et al. 1997). Conclusion
To conclude on current data, Kenyan tomato landraces are found to be suitable for production systems where processing is the commercial objective. Furthermore, due to their ability to produce a high quality of fruit over time, their usefulness for improving tomato production under intensive and continuous systems cannot be ignored. However, modern cultivars produce higher fruit yield and will remain equally important for tomato improvement. In addition, due to the ever increasing rate at which tomato is introduced, there is a need to develop a reliable, faster and affordable cultivar characterization procedure in order to safeguard small-scale producers. Acknowledgements
This work was accomplished with the help of a financial grant from the German Academic Exchange Service (DAAD) to S. G. Agong. The authors also wish to thank Mr. E. Sommer, Mr. B. Arnemann and Mrs C. Methner for their unfailing technical support. References
Copyright 2001 The Journal of Food Technology in Africa, Nairobi The following images related to this document are available:Photo images[ft01005t1a.jpg] [ft01005t1b.jpg] [ft01005f2.jpg] [ft01005t2.jpg] [ft01005f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}