 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Journal of Food Technology in Africa, Vol. 6, No. 3, July-Sept, 2001 pp. 96-100 Inter cropping and population density effects on yield component, seed quality and photosynthesis of sorghum and soybean
Elijah M. Akunda
Department of Botany University of Nairobi P.O. Box 30197, Nairobi-Kenya Code Number: ft01026
Abstract
Population density and intercropping of sorghum (sorghum bicolour (L) and Soybean {Glycine max Merril (L)} may alter their growth and yield patterns through responses to light and physiological attributes as compared to their performance in standard densities and monocultures. There is limited field information on this hypothesis. Thus the objective of this study was to determine the influence of intercropping and population density on protein and oil yield components, photosynthesis of sorghum and Soybean at the canopy closure. The study was conducted at the University of Nairobi farm during the long rains. There was a significant increase in the number of Soybean pods per plant between the low density and normal density populations followed by a decline at high plant populations. Whereas population density increased the protein seed yield of Soybean, intercropping hardly altered it. In contrast, both population density and intercropping significantly decreased Soybean seed oil content. The observed protein and oil content is explained by the photosynthetic responses of partial shaded and fully exposed leaves. There was a significant decrease and increase of Soybean photosynthetic rates of both non-shaded and partially shaded leaves respectively within the intercrops, whereas population density caused a significant decline in non-shaded and partially shaded leaves. The interactive effects between population density and intercropping caused an increase in percent Soybean seed protein and decreases in its percent oil content and photosynthetic rates of partially shaded sorghum and Soybean leaves. These results demonstrate that population density and intercropping may be a viable alternative of manipulating the productivity of these crops through their changes in physiological processes. Key words: Population density, intercropping, Soybean, sorghum, yield components, photosynthesis percent protein and oil yields. Introduction
The yield and quality of seeds are affected by the growth conditions of crops. Thus the amount of protein and oil content can be influenced by agronomic practices and growth characteristics of crops. Sexton and Shilber (1999) demonstrated that most of the sulphur containing amino acids synthesized during reproductive growth could be synthesized in the seed itself than importing it from other tissues. Yield is one of the most complex agronomic traits (Orf et al., 1999). It is mainly directed by various physiological process and is influenced by physical resources such as light, water and nutrients. Crop yields are further complicated by cultural practices such as intercropping and population density. Responses of crops to all the above resources are manifested through yield components, and physiological processes among others (Connor, 1983). For example, Soybean yield can be limited by the activity of the source or the ability of the seed to assimilate produced by the source (Egli, 1999).
The duration of Soybean seed filling is affected by resources such as water, (Korte et al., 1983), solar radiation, (Board and Harville 1996). Variation in environmental resources may bring about alterations in growth patterns of crops, for example leaf areas. Individual leaf areas depend on cell number and cell size while the duration of seed filling varies greatly according to field conditions. For example, Traore and Meranville (1999) demonstrated that in grain sorghum nitrate reductase activity was not a factor that limited nitrogen accumulation and use efficiency. On the other hand in indeterminate Soybean, determination of seed filling has been described as the result of the "self destruction" of the plant due to remobilisation of large amounts of nitrogen (Sinclair and de Wit, 1976).
Environmental conditions during the growth period, especially intensity and quality of solar radiation intercepted by the canopy, are important determinants of yield components, and photosynthesis. For example, shading (49 - 20%) of ambient light resulted in lengthening of internodes and increased lodging of Soybean plants (Ephrath et al., 1993). It has been demonstrated by Egli (1999) that the canopy photosynthesis during flowering and pod set of Soybean is an important determinant of seeds per meter square and that the impact of shading on seeds per metre depends on the duration of shading. The shading effects are more pronounced by cultural practices of intercropping and population density. Planting Soybean in narrow rows (<50cm) caused an increase in seed yield and this was attributed to increased light interception during vegetative and early reproductive periods (Wilcott et al., 1984). The increases in light interception can influence plant row yield, which is an important indicator of yield potential of the plant (Hegstad et al., 1999).
There is a differential response of yield components to changes in the environmental contribution. Also there are reports that show spacing affects percent oil content in seeds of crops (Hinson and Hanson 1962). Changes in growth conditions impacts on the yield components of crops. Pod number per plant is an example of an important yield component most influenced by alternations in cultural environments (Herbert and Litcufield, 1982). Similarly expansive growth of leaves is also influenced, which inturn affect the photosynthetic rates of crops. Any reductions in expansive growth of leaves is crucial to photosynthesis as it defines the size of its canopy for capturing sunlight (Hsiao, 2000). Photosynthesis of leaves have wide range of effects on the growth of crops. It influences the processes involved in carbon partitioning to various organs of a plant (Whisler et al., 1986). Usually, carbon is transported from a mature leaf to the nearest actively growing region, but this general pattern can be changed if leaves are removed or shaded (Sibles et al., 1987). Although growth patterns of crops are inherently affected by physical resources and cultural environments, there is little knowledge of the influences of population density and intercropping on physiological processes of field grown crops. Thus the objective of this study was to determine the influence of intercropping and population density on protein and oil yield of Soybean, yield components, photosynthesis, as partial and non shaded leaves of sorghum and Soybeans under field conditions. Materials and methods
Sorghum {Sorghum bicolour. (L)} cultivator Serena and Soybean {Glycine Max Merril (L)} cultivator Congo were raised in the field during the long rains (March to June) in the University of Nairobi farm. The two crops were planted at three population densities designated as pure and intercrops at a population ratio of 1:1 with a spacing of 75cm to 20cm; 60cm to 20cm; 45cm to 20cm. A split plot design was used with the density comprising the main plot while crop combinations forming subplots each replicated four times. Each subplot was planted with either monoculture and intercropped culture of the two crops and surrounded by 75 cm of bare earth.
The Soybean seeds were inoculated before planting with Rhixozium strain 7210 supplied by Nairobi MERCEN. Phosphate was supplied in the form of P2O5 at the rate of 45kg/ha at the planting time.
In this study, the growth measurements carried out included head length, number of branches, pods, stem diameter and photosynthetic rates. Sub samples of four plants were randomly taken at the centre of the plots for determination of the growth attributes.
Field photosynthetic rates were estimated using a field ADC intra-red gas analysis in an open system mode. This system also measured light intensity. The light intensity was measured by a quantum sensor mounted on the leaf chamber sampler. Photosynthetic and light measurement were taken on a relatively sky clear days with a minimum overcast. A known area of the leaf was enclosed within a small chamber, which was adequately ventilated by a miniature fan. This system also monitored air temperature and relative humidity. Photosynthetic measured were taken on partially and non-shaded leaves of Soybean and sorghum. The rates of photosynthesis were obtained by calculation employing Kumar and Tieszen, 1980 method.
The percentage seed pattern content was determined from total nitrogen. This was obtained by multiplying the total percent nitrogen with a factor of 6.25%. The oil content was soxhlet extracted using n-hexane as the solvent for 12 hours. Dehydration was done by using magnesium sulphate then followed by filtration and evaporative removal of the solvent in vacuo. The contents in the flask were subjected to further evacuation overnight with concomitant heating in warm water to remove any laboratory traces of hexane before final weight was determined. In order to establish the efficiency and purity of the extracted oil, refractive indices of the samples were determined. Results
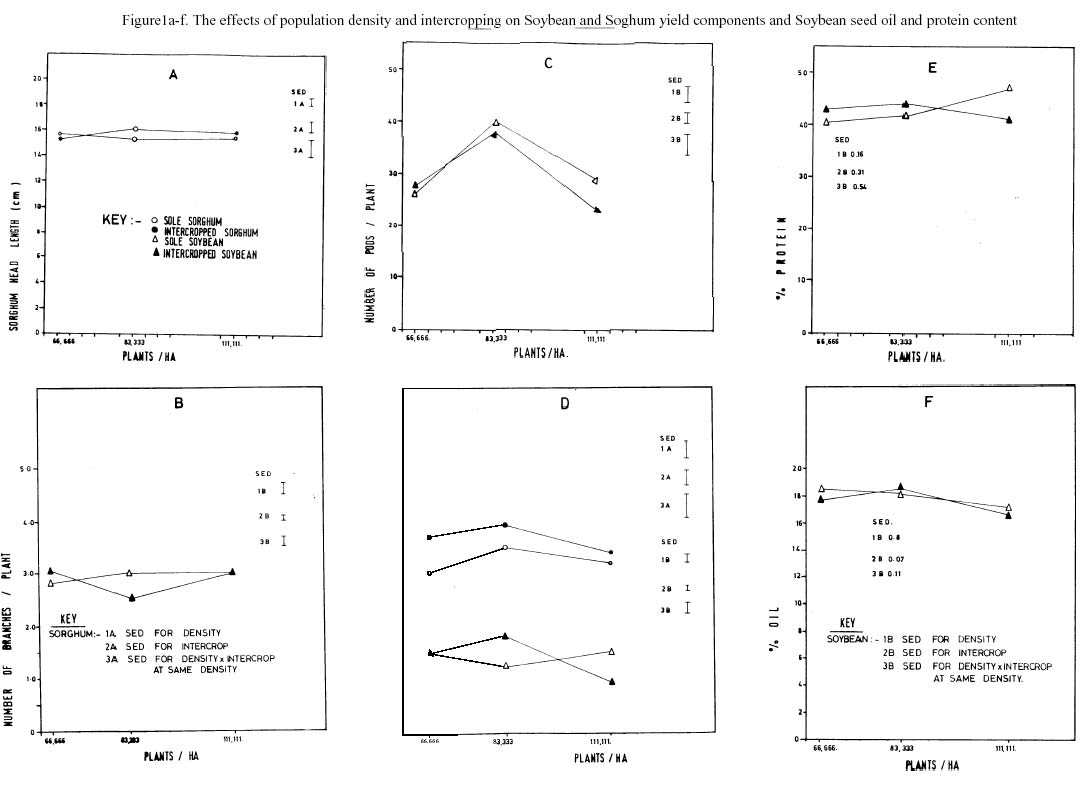
The effects of population density and intercropping on Soybean and sorghum yield components and Soybean seed oil and protein content is shown in figure 1a, b, c, d, e & f. Intercropping and population density did not influence sorghum head length (fig. 1a). The observed decrease and increase in primary Soybean and within intercrops and with increasing population density were not significant (fig 1b). However, the number of pods decreased non-significantly within intercrops but significantly (p = 0.01) caused an increase at normal density and a decline at high density stands of Soybean. Both the effects of intercropping and population density were insignificant for sorghum and Soybean stem diameters. The interactive effects of population density and intercropping were noticed only for Soybean. There was a significant decreasing interactive effect of intercropping and population density on branch numbers, and pod numbers (p = 0.01).
The percent seed protein yield of Soybean was hardly influenced by intercropping, the small decrease within intercrops was however non significant (Fig 1e). Population density influenced Soybean seed protein content. There was a highly significant p = 0.01) increase of seed protein content at high plant density. Whereas the interaction of population density and intercropping caused a significant increase in percent protein, there was a significant (p = 0.01) decrease for percent oil content. The light intensities of non shaded leaves ranged from 1600 to 1700 mmol m-25-1 and 14000 and 1500 mmol m-25-1 in the morning and afternoon respectively, in contrast to partial shaded leaves whose light intensities ranged from 60 to 83 mmol m-25-1 and 60 to 70 mmol m-25-1 in the morning and afternoon respectively.
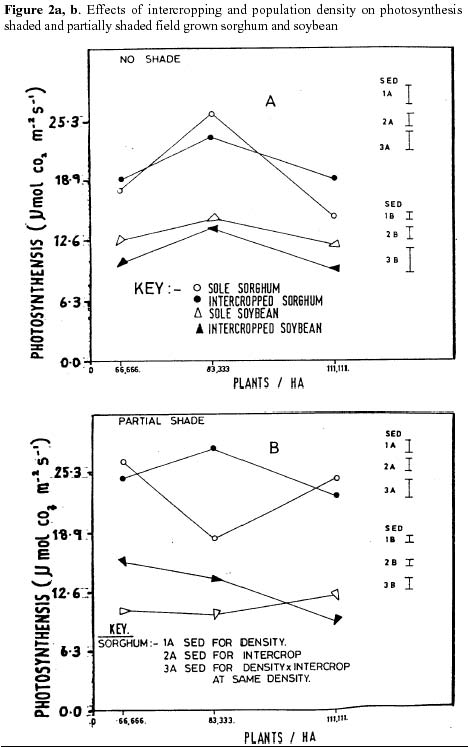
Figure 2a and b represent the photosynthetic rates of Soybean and sorghum under no shade and partial shade under field conditions respectively. Although intercropping hardly influenced the photosynthetic rates of sorghum of both partially shaded and non shaded leaves there was a significant influence of density on non shaded leaves. The increase in photosynthesis at normal density followed by a decrease at high density followed by a decrease at high density was significant (p = 0.05). The decrease in photosynthetic rates of sorghum leaves with increase in population density for partially shaded leaves were non significant (fig. 2b).
The decrease in intercropped Soybean photosynthetic rates of non shaded leaves and the increase in partially shaded leaves were significant (p = 0.05). Population density caused a significant (p = 0.05) decreasing effect of photosynthesis on both non shaded and partially shaded Soybean leaves. Both sorghum and Soybean partially shaded leaves exhibited a significant photosynthetic decreasing effect of the interaction between effect of the interaction between population density and intercropping (Fig. 2b). Discussion
The seed oil of Soybean decreased (37%) significantly in the intercropped stands. Intercropping sorghum and Soybean influences Soybean quality attributes. Redfearn et al., (1999) similarly noted that intercropping of sorghum and Soybean affected the yield, morphology and quality of forage Soybean. The observed percent oil content of Soybean seeds in the intercrops indicated a competition for resources between Soybean and sorghum. In the sorghum and Soybean intercrops, the sorghum is the dominant crop. Its aggressiveness thus influences the distribution of environmental resources of light, water and nutrients in such combination, there is therefore in equitable distribution of growth resources. It is likely that the decline in oil content was due to the Soybean having been dominated by the sorghum. The dominating effect of sorghum is crucial during the reproductive stage of Soybean. This may inturn have influenced the quality attributes such as the percent oil content. Lesoing and Francis (1999) noted that competition for resources in Soybean, maize and grain sorghum grown in strips was important in both reproductive and grain filling periods.
Increasing population density of Soybean significantly increased protein content by 57%. This is indicative of population density favouring nitrogen assimilation. In order to assimilate large quantities of nitrogen, there should be high leaf crude protein concentration, adequate photosynthesis (Bruening and Egli, 1999), increased stem diameter, number of branches and seed growth. For example Redfearn et al., (1999) demonstrated that stems from intercropped Soybean had greater crude protein than mono cropped stems.
Population density and intercropping are major cultural practices that impact on light regimes of the canopy. Thus the observed trends in photosynthetic rates of non shaded and partially shaded leaves of sorghum and Soybean leaves were influenced by the changes in solar interception. This is evidenced by the significant decreases and increases of Soybean photosynthetic rates of non shaded and partially shaded respectively in the intercrops and the observed decreased rates of partially shaded and non shaded leaves with increasing population density. Population differences will influence timing and severity of interplant competition, and affect canopy structure and efficiency. For example in some Soybean cultivators, planting density may influence the extent of fibrous like root system, which contributes to enhanced drought tolerance (Pantalone et al., 1999). Such competitive effects may explain the observed photosynthetic decreases of sorghum in non shaded leaves. Egli (1999) observed similar results in Soybean, in which shade almost significantly reduced the individual seed growth of Soybean.
The foliage arrangement has been shown by Fischer and Wilson (1976) to be strongly influenced by population density. Thus an increase in population density would imply a corresponding increase in light interception. In cases where this was not the case as indicated above, this showed that to better understand how crops intercept light, the complexity of plant structure needs to be characterised. The observed declines in photosynthetic rates of non shaded leaves with population increase may partially be explained by leaf area. Light absorption depends on rate of leaf expansion (Hsiao, 2000), leaf disposition and time of senescence (Bruening and Egli, 1999). Light penetration decreases during canopy development and this is further complicated by population density and intercropping. Similar results were noted by Foroutan-pour et al., (1999) in which he demonstrated that Soybean canopy development was affected by population density and intercropping.
On the overall this study shows that careful understanding of yield components, and photosynthetic performance can add to the comprehension of crop performance. Also by understanding the effects of population density and intercropping with respect to physiological attributes such as percent oil and protein content, photosynthesis can contribute to knowledge of designing productive systems. References
Copyright 2001 The Journal of Food Technology in Africa, Nairobi The following images related to this document are available:Photo images[ft01026f1.jpg] [ft01026f2.jpg] |
| |||||||||

{kind=link}
{kind=link}