 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Journal of Food Technology in Africa Vol. 7 No. 3, 2002, pp. 93-97 Identification of Lactic Acid Bacteria isolated from Opaque beer (Chibuku) for potential use as a starter culture Chamunorwa A.Togo1*, Sara B. Feresu2, Anthony N. Mutukumira3 1) Department of Biological Sciences, Midlands State
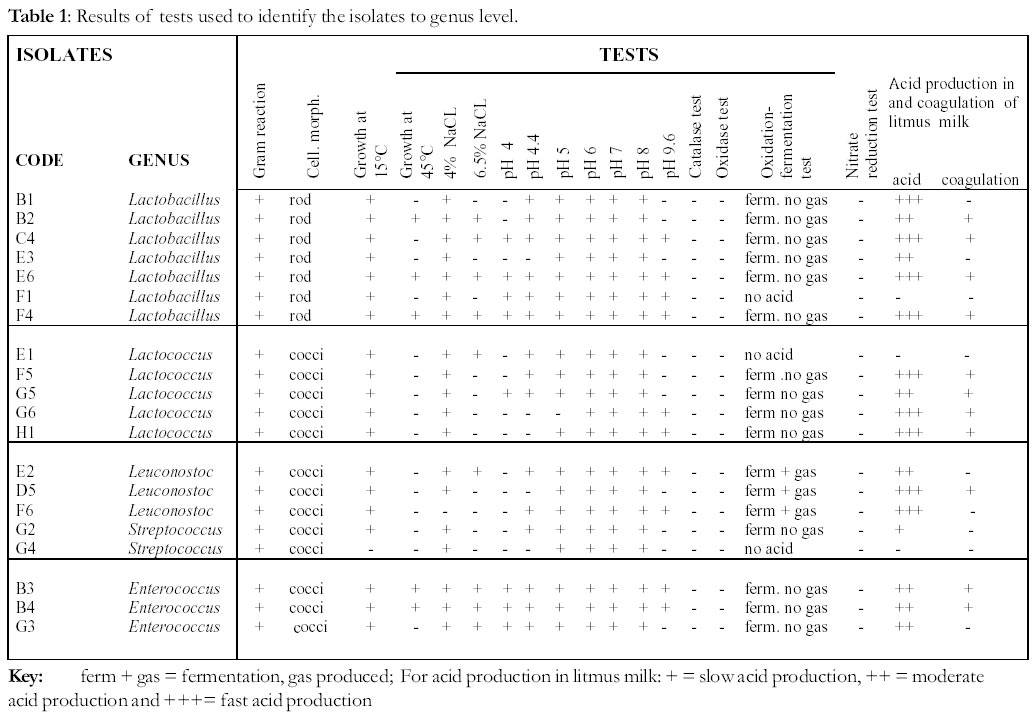
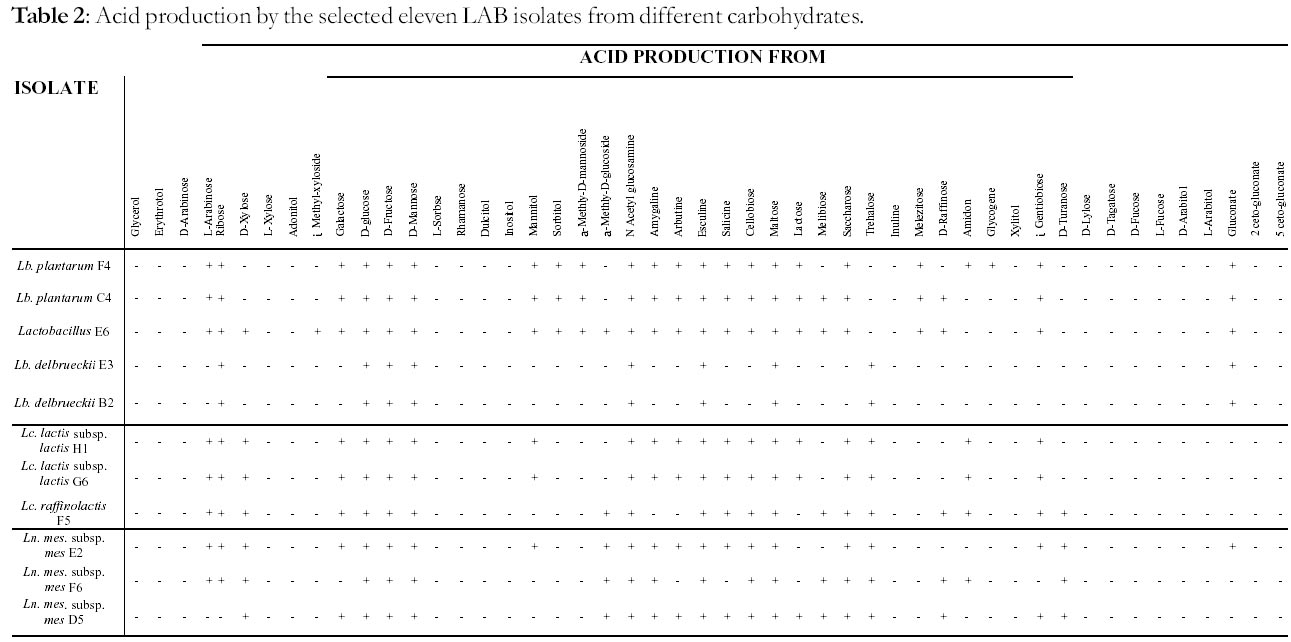
University, P. Bag 9055, Gweru, Zimbabwe *Corresponding author Code Number: ft02022 ABSTRACT A study was carried out to identify lactic acid bacteria (LAB) isolated from chibuku that would be later assessed for potential as starter cultures. Thirty-eight isolates were Gram stained and the 20, which were Gram positive, were identified to genus level using morphological, physiological and biochemical tests. Five genera of lactic acid bacteria were identified and these were Lactobacillus (seven isolates coded B1, B2, C4, E3, E6, F1 and F4), Lactococcus (five isolates: E1, F5, G5, G6 and H1), Leuconostoc (three isolates: E2, D5, F6), Streptococcus (two isolates: G2 and G4) and Enterococcus (three isolates: B3, B4 and G3). From these genera, eleven isolates five from the genus Lactobacillus, three from Lactococcus and three from Leuconostoc were selected for identification to species level using API 50 CH kits. The Lactobacillus strains were identified as follows: two strains were Lb plantarum (C4 and F4), two strains Lb. delbrueckii (B2 and E3), one strain could not be assigned to a species and was termed Lactobacillus sp. E6. Two of the Lactococcus isolates were identified as Lc. lactis subsp. lactis (G6 and H1), while the third isolate was Lc. raffinolactis (strain F5). The three Leuconostoc strains were Ln. mesenteroides subsp. mesenteroides. Key words: Lactic acid bacteria; Sorghum beer (chibuku); Starter culture; Identification INTRODUCTION There are two types of beer produced at an industrial scale in Zimbabwe; clear and sorghum/opaque beer. Clear beer is a product of alcoholic fermentation of barley malt and maize adjuncts by a bottom fermenting yeast Saccharomyces carlsbergensis (Casida, 1987; Varnam and Sutherland, 1994). Sorghum beer is a product of two fermentations of sorghum malt and straight run maize meal, an alcoholic fermentation by a top fermenting yeast Saccharomyces cerevisiae and a spontaneous lactic acid fermentation (Novellie, 1986; Haggblade and Holzapfel, 1989; Steinkraus, 1996). The lactic acid fermentation in sorghum beer is effected by microorganisms inherent in the raw materials, containers and the surrounding environment (Marshall, 1987; Tamime, 1990; Nout, 1992; Mpandi-Khosa, 1993). Spontaneous fermentations are difficult to control; are not predictable in terms of length of fermentation and quality of product; can produce unwanted products or products with a short shelf life and are sometimes not safe since they are liable to contamination by pathogens (Novellie and De Schaeprijver, 1986; Tamime, 1990; Nout, 1992). To overcome this problem, the most predominant microorganisms found in an acceptable product are isolated and purified (Marshall, 1987; Tamime, 1990; Marshall, 1993). The medium used for the fermentation is then pasteurized to exclude most unknown microorganisms and the purified microorganism(s) is/are introduced to initiate the fermentation (Marshall, 1987; Hesseltine, 1992; Marshall, 1993). By so doing, the fermentation can be manipulated in such a way that it is possible to predict the amount and quality of product formed, and the length of the fermentation period (Tamime, 1990; Hesseltine, 1992). Such introduced cultures are termed starter cultures (Hesseltine, 1992; Marshall, 1993; M‰yr‰-M‰kinen and Bigret, 1993). The most commonly sold sorghum beer in Zimbabwe is termed chibuku. Chibuku has inherent problems of poor and inconsistent quality. Although commercially prepared lactic acid is added during the preparation of wort for chibuku brewing, spontaneous lactic acid fermentation (due mostly to mesophilic lactic acid bacteria inherent in the malt) has been demonstrated during the later stages of chibuku brewing (Mashanda, 1997). This spontaneous lactic acid fermentation might be one of the contributing factors to the inconsistent quality of chibuku (Novellie and De Schaeprijver, 1986; Gadaga et al., 1999). Thus the main objective of this study was to purify and identify the most predominant LAB that had been isolated from chibuku for later assessment of their potential as lactic acid bacteria starter cultures. The identified genera and species were reportedly associated with sorghum grains and some were found to occur in South African sorghum beers. In Zimbabwe, no work of this nature has been carried out on either chibuku or any sorghum beer. Instead, the only related work was on the determination of shelf life of chibuku that was found to be five days (Mashanda, 1997). MATERIALS AND METHODS Source, purification and maintenance of isolates The isolates were obtained from the Department of Biological Sciences, University of Zimbabwe and had been isolated from 1-5 day old chibuku using MRS agar (Oxoid) and stored at 4±1ºC. The purity of the isolates had not been previously authenticated. Morphologically different colonies were picked from the plates and each streaked on a separate plate of MRS agar (Biolab). The plates were then incubated at 30±1ºC for 3 days and the resulting colonies examined for purity. The process was repeated until there were no mixed cultures on each plate. Thirtyeight morphologically different isolates were obtained for further investigations. Each isolate was kept in undiluted glycerol at -80°C as a means of long maintenance (Lapage and Redway, 1973). Working cultures were maintained on plates kept at 4±1°C with fortnight subculturing on MRS agar (Collins and Lyne, 1987). Identification of isolates to genus level The isolates were identified to genus level by Gram staining (Harrigan and McCance, 1993), observing growth at 15 and 45°C, different temperatures and growth in saline MRS broth(4 and 6.5 % NaCl w/v). Other tests were catalase test (Harrigan and McCance, 1993), oxidase test (Collins and Lyne, 1987), nitrate reductase test, oxidationfermentation test and growth in litmus milk (Harrigan and McCance, 1993) The results obtained for morphological, physiological and biochemical tests were compared with those in standard texts for identification (Sharpe, 1981; Teuber and Geis, 1981; Garvie, 1986; Kandler and Wiess, 1986; Mundt, 1986; Dellaglio et al., 1995; Devriese and Pot , 1995; Hammes and Vogel, 1995; Hardie and Whiley, 1995; Teuber, 1995) and the isolates assigned to appropriate genera. Identification of selected isolates to species level using API 50 CH system Eleven isolates belonging to the genera Lactobacillus (5), Lactococcus (3) and Leuconostoc (3) selected based on the rate of acid production in litmus milk were identified to species level using API 50 CH kits and L media (BiomÈrieux, France) according the manufacturer's instructions. The API profiles were analysed using API LAB Plus (BiomÈrieux, France). In cases of equivocal results, reference was made to standard texts for identification. RESULTS Identification of the isolates to genus level Of the 38 purified isolates, 20 were Gram positive bacteria and were presumed to be LAB while the rest were either yeasts (10) or Gram negative members of the family Enterobacteriaceae (8). All the 20 Gram positive bacteria were negative for the catalase, oxidase and nitrate reduction tests (Table 1). The majority of the isolates were identified as belonging to the genus Lactobaccillus (7) with the rest belonging to the genera Lactococcus (5), Leuconostoc (3), Enterococcus (3) and Streptococcus (2) (Table 1). All members of the genus Lactobacillus (strains B1, B2, C4, E3, E6, F1 and F4) grew at 15°C, in 4% NaCl and at pH 5, 6, 7 and 8 (Table 1). Except for strain F1, all the other strains were fermentative with no gas production and produced acid in litmus milk (Table 1). The strains B1, C4, E3 and F1 did not grow at 45°C, while strains B1, B2 and E3 did not grow at pH 4 and 9.6. Strain E3 was the only one that could not grow at pH 4.4. Strains B1, E3 and F1 did not grow at 6.5% NaCl neither did they coagulate litmus milk (Table 1). All strains belonging to the genus Lactococcus (strains E1, F5, G6 and H1) had the ability to grow at 15°C but not 45°C, in 4% NaCl, and at pH 6, 7 and 8 (Table 1). Strain E1 reacted differently from other Lactococcus strains by growing in 6.5% NaCl, not fermenting glucose, not producing acid in litmus milk and not coagulating the litmus milk (Table 1). Isolate G5 was the only strain which could grow at pH 4 while isolate G6 was the only strain which showed no growth at pH 5. Isolates E1 and F5 grew at pH 4.4 while H1 could not grow. Only strains H1 and G6 could grow at pH 9.6 (Table 1). The three Leuconostoc strains (E2, D5 and F6) grew at 15°C but not at 45°C, grew at pH 5, 6, 7 and 8 but not pH 4, were fermentative with gas production and produced acid from litmus milk (Table 1). Strains E2 and F6 could grow at pH 4.4 and 9.6 but could not coagulate litmus milk while strain D5 showed opposite reactions for all the three tests (Table 1). Only strain E2 could grow in 6.5% NaCl while only strain F6 could not grow at 4% NaCl (Table 1). The two strains of Streptococcus (G2 and G4) showed growth in 4% NaCl and at pH 5, 6, 7 and 8 but which did not grow at 45°C, in 6.5% NaCl and at pH 4 and 9.6, and which could not coagulate litmus milk (Table 1). Strain G2 was more reactive than strain G4 as it could grow at 15°C and at pH 4.4; could ferment glucose but with no gas production and could produce acid from litmus milk all which were negative for strain G4 (Table 1). The three strains belonging to Enterococcus (B3, B4 and G3) had the ability to grow at 15°C, in 4% and 6.5% NaCl and at pH 4, 4.4, 5, 6, 7 and 8; were fermentative with no gas production and could produce acid from litmus milk (Table 1). Strain G3 differed from other two isolates by not being able to grow at 45°C and pH 9.6 and not coagulating litmus milk (Table 1). Identification of selected isolates to species level Five strains of Lactobacillus (C4, F4, E6, E3 and B2), three each of the genus Lactococcus (H1, G6 and F5) and Leuconostoc (E2, F6 and D5) were identified to species level. Two Lactobacillus strains (C4 and F4) were identified as Lb. plantarum, strains B2 and E3 were identified as Lb. delbruekii while strain E6 could not be assigned to a species (Table 2). Although strains C4 and F4 were identified as Lb. plantarum they differed in their ability to ferment melibiose, D-raffinose, amidon and glycogene (Table 2). Lactobacillus sp. (E6) was capable of metabolizing the highest number of sugars (26 out of 49) while the two Lb. delbrueckii only metabolized nine sugars (Table 2). Two Lactococcus strains (H1 and G6) were identified as Lc. lactic subsp. lactis and they had identical biochemical profiles (Table 2). The third Lactococcus strain F5 was identified as Lc. raffinolactis. The Lc. lactis subsp. lactis strains could be distinguished from the Lc. raffinolactis strains on the basis of their acid production from eight carbohydrates (Table 2). The three Leuconostoc strains (E2, F6 and D5) were identified as Ln. mesenteroides subsp. mesenteroides. However, their acid production from ten carbohydrates was variable (Table 2). DISCUSSION Lactic acid bacteria have been consistently demonstrated to be responsible for the spontaneous lactic acid fermentation of sorghum beer (Novellie and De Schaeprijver, 1986; Haggblade and Holzapfel, 1989; Steinkraus, 1996). This study has confirmed the presence of LAB in chibuku as five genera of LAB, Lactobacillus, Lactococcus, Leuconostoc, Streptococcus and Enterococcus, were predominant in chibuku. With the exception of the genus Enterococcus, the other genera have been previously reported in African sorghum beers (Novellie and De Schaeprijver, 1986; Haggblade and Holzapfel, 1989). Similar to observations in South African beers, the most predominant LAB in chibuku were members of the genus Lactobacillus while leuconostocs were also present but in lower numbers (Novellie and De Schaeprijver, 1986; Haggblade and Holzapfel, 1989). The lower numbers of leuconostocs is probably due to their inability to compete with other LAB in mixed cultures (Teuber and Geis, 1981). The isolation of Lactococcus strains from chibuku is consistent with their previously reported presence in sorghum malt (Teuber, 1995). The least number of isolates from chibuku belonged to the genus Streptococcus similar to previous observations in other sorghum beers. The source of the Enterococcus strains is not clear since they have not been previously reported in sorghum beer. However, members of this genus are commonly found in plant material (Devriese and Pot , 1995) and therefore they could have been introduced by the sorghum malt or maize. When selected strains of the genus Lactobacillus were identified to species level as Lb. plantarum and Lb. delbrueckii, one strain could not be allocated to a species based on the API 50 CH tests since the strain's profile was judged equally as Lb. plantarum and Lb. pentosus. This was therefore designated Lactobacillus sp. (E6). Differentiation of Lb. plantarum and Lb. pentosus by the tests used is difficult as these two species behave similarly (Kandler and Wiess, 1986). However, ribosomal RNA analysis can be employed to differentiate the two. The two species of lactobacilli identified in chibuku have also been reported in other sorghum beers. Similar to observations made in this study, Lb. plantarum has been identified as the dominant species in sorghum beers while Lb. delbrueckii occurs in low numbers and can sometimes be absent (Novellie and De Schaeprijver, 1986; Haggblade and Holzapfel, 1989). However, the occurrence of lower numbers of Lb. delbrueckii in chibuku could be due to the isolation temperatures (30°C) used that favoured growth of mesophiles like Lb. plantarum while selecting them against growth of thermophiles such as Lb. delbrueckii (Hammes and Vogel, 1995). The strains belonging to the genus Lactococcus were identified as Lc. lactis subsp. lactis and Lc. raffinolactis. Lactococcus lactis has been reported in South African sorghum beers (Haggblade and Holzapfel, 1989) while this is the first report of Lc. raffinolactis in such beers. Reports of occurrence of Lc. raffinolactis on plant material (Teuber, 1995) helps to explain their presence in sorghum beer. Leuconostoc mesenteroides subsp. mesenteroides which were identified in chibuku have been previously reported as spoilage microorganisms in sorghum beers due to the excessive levels of organic acids and volatile compounds they produce (Novellie and De Schaeprijver, 1986; Haggblade and Holzapfel, 1989). Their role as spoilage organisms has however not been substantiated since sorghum beer is a live product and even the presence of desirable LAB sometimes results in an undesirable product due to over production of such organic acids and volatile compounds. The identified isolates will undergo tests for lactic acid production and selected for further tests (production of desirable organic acids and volatile compounds) to assess their potential as starter culture in the brewing of sorghum beer. The lactic acid starter culture will greatly contribute in solving the problem of inconsistent quality and short shelf life of sorghum beers in Zimbabwe as the fermentation process will be under full control. ACKNOWLEDGEMENTS The authors are grateful to Sibonginkosi Moyo for isolating the microorganisms, Henry Gadaga for his advice, Chibuku brewery for supplying the sorghum beer and the DGIS for funding this study. REFERENCES
Copyright 2002 The Journal of Food Technology in Africa, Nairobi The following images related to this document are available:Photo images[ft02022t1.jpg] [ft02022t2.jpg] |
| |||||||||

{kind=link}
{kind=link}