 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
European Journal of General Medicine, Vol. 1, No. 3, 2004, pp. 26-32 ORIGINAL ARTICLE DOES THE SUBACUTE (4-WEEK) EXPOSURE TO FORMALDEHYDE INHALATION LEAD TO OXIDANT/ANTIOXIDANT IMBALANCE IN RAT LIVER? Sadık Söğüt1, Ahmet Songur2, Oğuz Aslan Özen2, Hüseyin Özyurt3, Mustafa Sarsılmaz4 1Mustafa Kemal University, Faculty of Medicine, Department of Biochemistry, 2Afyon
Kocatepe University, Faculty of Medicine, Department of Anatomy, 3Gaziosmanpaşa
University, Faculty of Medicine, Department of Biochemistry, 4Fırat
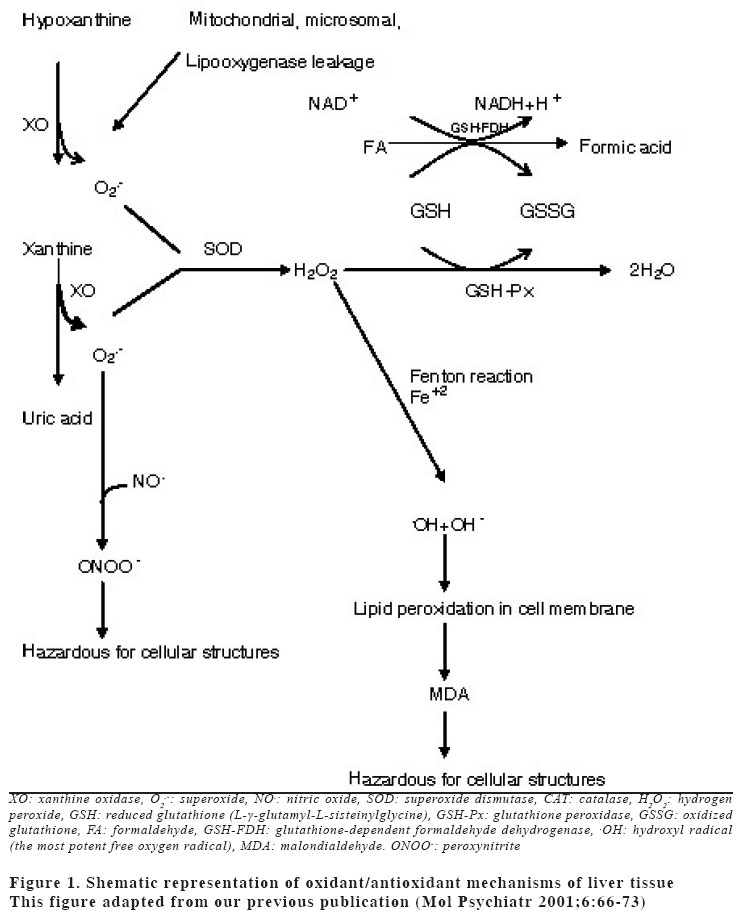
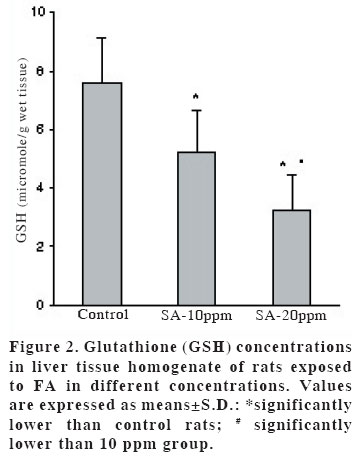
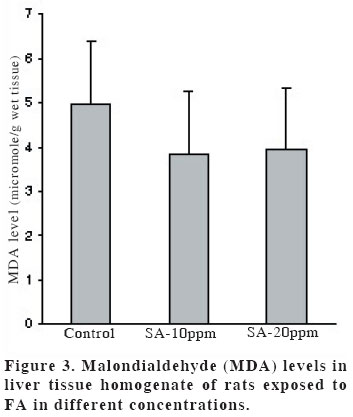
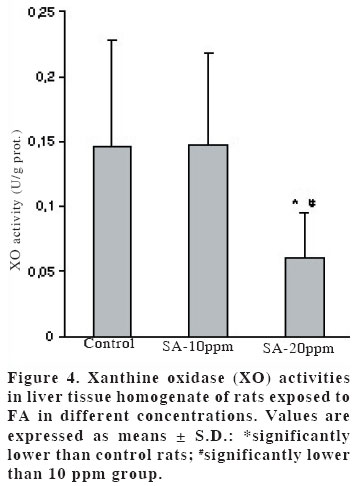
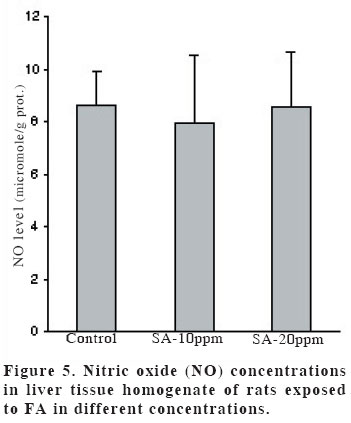
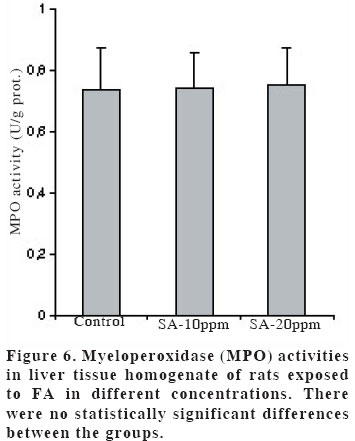
University, Faculty of Medicine, Department of Anatomy Code Number: gm04028 The aim of this experimental study is to evaluate oxidant/antioxidant status and the end products of lipid peroxidation in the liver of rats exposed to subacut formaldehyde (FA) inhalation (4-weeks). Thirty male Wistar albino rats divided into 3 separate groups randomly (10 for each group). Rats were exposed to atmospheres containing 0, 10 and 20 ppm FA continuously (8 hours per day, 5 days per week). Reduced glutathione (GSH), malondialdehyde (MDA) and nitric oxide (NO) levels, as well as xanthine oxidase (XO), and myeloperoxidase (MPO) activities were measured in rat liver homegenates. The results showed a remarkable reduction of GSH content (10 and 20 ppm FA) and XO activity (20 ppm FA) in liver tissues from rats exposed to for 4weeks (p<0.005, p<0.0001 and p<0.0001, respectively). MDA and NO levels did not change in both two groups (10 ppm and 20 ppm). These findings suggest that the antioxidant system of liver tissue is moderately impaired by excessive FA exposure. The GSH has an important influence especially on subacute FA liver toxicity. GSH-related cellular defensive mechanisms may be deppressed and susceptibility to oxidative damage may high in rats exposed to FA subacutely. Key words: Formaldehyde, liver, lipid peroxidation, glutathione, oxidant/antioxidant enzymes INTRODUCTION Formaldehyde (FA) has been used extensively to protect cadavers from decaying for decades in Anatomy laboratories of Medical Faculties in Turkey (1). It is obviously known that it has some adverse effects on human body. It has been also shown that FA may reveal certain pathologies: When it is ingested orally, plasma protein and albumin levels decreased, hyperkeratosis in the forestomach, and focal gastritis in the glandular stomach appeared (2). All types of gene mutations and chromosomal rearrangements were reported (3) and the damaged chromosome cells were 43% as compared to 1,7% of control value (4). Formaldehyde was found to induce rhinitis, degeneration, frank necrosis, hyperplasia and squamous metaplasia of the ciliated and non-cliated nasal respiratory epithelium (5). Feron et al. reported that when rats exposed to FA by short-term inhalation nasal respiratory epithelium severely damaged by FA vapour often does not regenerate and in some cases tumors develop (6). It is still obscure whether FA can induce toxic effects in distant organs such as liver, kidney, testes, etc. Glutathione-dependent formaldehyde dehydrogenase (GSH-FDH) has an universal role in biological FA oxidation to formic acid (7) (Figure 1). We hypothesized that when FA was ingested and/or inhaled excessively, GSH-FDH will detoxify this compound more than normal metabolic situation and GSH will run out in the liver which is the main metabolic organ in the human body. GSH (L-g-glutamyl-L-cysteinylglycine) has antioxidant properties and is abudantly present in liver tissue. GSH is involved in the reactions by various enzymes and it protects against reactive oxygen species (ROS) and toxic compounds such as FA. Xanthine oxidase (XO) is one of the major in vivo superoxide generator and activated to utilize accumulated purines in the ischemic part of the organ in some circumstances such as ischemia/reperfusion. It is very important to know whether XO activity of the liver tissue is increased or decreased, because oxygen radicals and antioxidants play an important role in the detoxifying process of FA. The aim of this study is to evaluate antioxidant status and lipid peroxidation in the liver of rats exposed to FA by inhalation subacutely (4-weeks). For this purpose, GSH as an index of non-enzymatic antioxidant status of the liver tissue; malondialdehyde (MDA), as an index of in vivo lipid peroxidation; xanthine oxidase, a superoxide radical (O2.-) generator; nitric oxide (.NO), a free radical; and myeloperoxidase (MPO), as an index of leukocyte sequestration to the liver tissue were studied in liver homogenates. MATERIALS AND METHODS Chemicals and instruments The production of FA gas Animals and procedure of FA exposure The preparation of liver tissues for biochemical analyses MDA level determination Following the reaction, fluorescence intensity was measured in the n-butanol phase with fluorescence spectrophotometer adjusting wavelengths excitation at 525nm and emission at 547nm, by comparing with a standard solution of 1,1,3,3 tetramethoxypropane. Results were expressed in terms of µmol/g wet tissue. XO activity determination The reaction was stopped by addition of 0.1 mL 100% (w/v) TCA and the mixture was centrifuged at 5000 x g for 15 min. With resultant clear supernatant, absorption at 293 nm was measured against blank. One unit of activity was defined as 1 µmol of uric acid formed per minute at 37 °C, pH 7.5, and expressed in U/g protein. NO level determination GSH level determination MPO activity determination Statistical analysis RESULTS Figure 2 shows the effects of inhaled FA on liver GSH content. GSH levels were gradually decreased in both 10 ppm group (5.23±1.44 µmol/gr wet tissue) and 20 ppm group (3.24±1.21 µmol/gr wet tissue) when compared to control group (7.58±1.54 µmol/gr wet tissue) (p<0.005 and p<0.0001, respectively). FA was found to cause GSH depression in a dose-dependent manner. Figure 3 shows the effects of FA on liver peroxidation in rats subjected to subchronic FA inhalation. There was no statistically significant increase in tissue MDA level in 10 ppm and 20 ppm groups compared to both control group. The activity of liver oxidant enzyme XO is reported in Figure 4 for the two study groups. Rats exposed to FA resulted in a dose-independent inactivation of XO more than two-fold. There was a statistically significant decrease in XO activity in 20 ppm group compared to both control and 10 ppm groups. The level of the liver NO is reported in Figure 5. The results show that FA exposure is not associated with the changed NO production in both 10 ppm and 20 ppm exposure. The activity of liver MPO is presented in Figure 6. There were no activity changes of MPO in 10 ppm and 20 ppm groups compared to control group. There is a statistically significant correlation between NO and MDA in 20 ppm group (r= 0.676, p<0.035). DISCUSSION Free oxygen radicals are reactive oxygen species and reduction of molecular oxygen produces these radicals. Many of the toxic effects of oxygen can be attributed to these reactive oxygen species. Exposure of membrane lipids of liver cells to oxygen radicals in the presence of iron salts stimulates the process of lipid peroxidation. Liver GSH plays an important role in protecting membrane lipids of the liver cells from reactive oxygen species (ROS) attack (14); however it is rapidly oxidized when challenged by oxidants released from metabolic events or degrading processes such as GSH-FDH (Figure 1). This rapid depletion may be the reason for the low GSH values in liver tissue. We observed 31 % and 57 % decrease in GSH level of liver tissue in Group I and Group II, respectively. We suggest that this decrease might probably be in response to the extra production of ROS or excessive exposure to FA as stated by some authors (1517). Glutathione peroxidase (GSH-Px), which is present widely in liver tissue, needs GSH to detoxify hydrogen peroxide (H2O2). If the enzymatic antioxidant system can not work properly in liver tissues, there will be an extra free oxygen radical load. In the end of the process, cellular and organelle membranes, which are more sensitive to oxidative attacks than other cellular components, suffer from successive lipid peroxidation and led to be destroyed of cellular structures completely. Although it was not statistically significant, MDA levels were found to be lower in Group I and Group II than those of control animals. We suggest two probable explanations which are important for this contrary results: i) enzymatic and nonenzymatic defense systems against free oxygen radicals, except GSH, might be increased and thus, compansated the effect of GSH and GSH-dependent enzymes such as GSH-Px, glutathione reductase (GSH-Red) and glutathione-S-transferase (GSH-ST), ii) MDA released by the lipid peroxidation in liver tissues might be metabolized immediately by a mitochondrial MDA-metabolizing enzyme (low specific aldehyde dehydrogenase) and increase in MDA level is not seen. It was found that the content of GSH was significantly decreased in rat liver after methanol intoxication for 6 hours to 5 days (18-19). Methanol is oxidized by liver alcohol dehydrogenase to FA (20). Formaldehyde in turn is rapidly oxidized by GSH-FDH to formic acid. These findings seem to be concordance with our results. Taken together, one of the reasons of decreased GSH in liver tissue may be this enzymatic degradation process. It is very difficult to explain why XO activity decreases in liver tissues of rats exposed to FA in Group II. NO is a free radical with “double edged”pathophysiological role in liver tissue, because it can be toxic or protective (21,22). NO is thought to be toxic through its reaction product with O2 ·- the peroxynitrite anion (ONOO-), which can further decompose into NO2 and .OH (23). NO is synthesized by three isoforms of nitric oxide synthases (NOS). Neuronal and endothelial isoforms are regulated by intracellular Ca++. Metabolic reactions that could produce ROS are catalyzed by Ca++-activated enzymes: phospholipase A2 and NOS-1. Inducible NOS activity is independent of Ca++, and the enzyme is expressed in various cells, such as macrophages and neutrophiles in response to inflammatory stimuli. In our study, detection of normal NO levels in the study groups I and II suggested that overproduction of free radicals by NOS1 isoform had not been occured. In conclusion, GSH improves endurance performance and prevents liver lipid peroxidation during subacute FA inhalation in rats. The GSH content of liver tissue was affected from FA toxicity with dose-dependent manner in our study. Therefore, liver GSH has an important influence on subacute FA toxicity. GSH-related cellular defensive mechanisms may be deppressed in liver tissues and therefore susceptibility to oxidative damage may increase in rats exposed to FA subacutely. REFERENCES
Copyright 2004 - Medical Investigations Society The following images related to this document are available:Photo images[gm04028f6.jpg] [gm04028f4.jpg] [gm04028f2.jpg] [gm04028f5.jpg] [gm04028f3.jpg] [gm04028f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}