 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
European Journal of General Medicine, Vol. 2, No. 1, 2005, pp. 1-4 GnRH AGONIST STIMULATES OOCYTE NUMBER AND MATURATION IN MICE SUPEROVULATED WITH eCG AND hCG Ömer Coşkun, Mehmet Kanter Zonguldak Karaelmas University, Faculty of Medicine, Department of Histology
and Embryology, Zonguldak, Turkey Aim: This study was designed to investigate the effects
of gonadotropin-releasing hormone agonist (GnRHa) on the ovulation rate, the
oocyte number and maturation in mice superovulated with equine chorionic gonadotropin
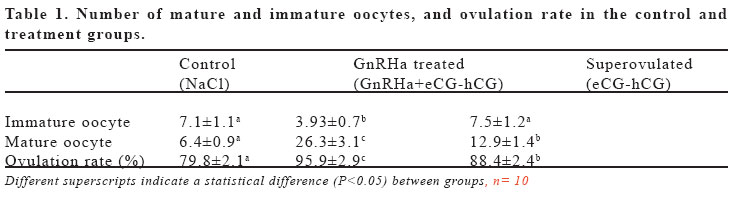
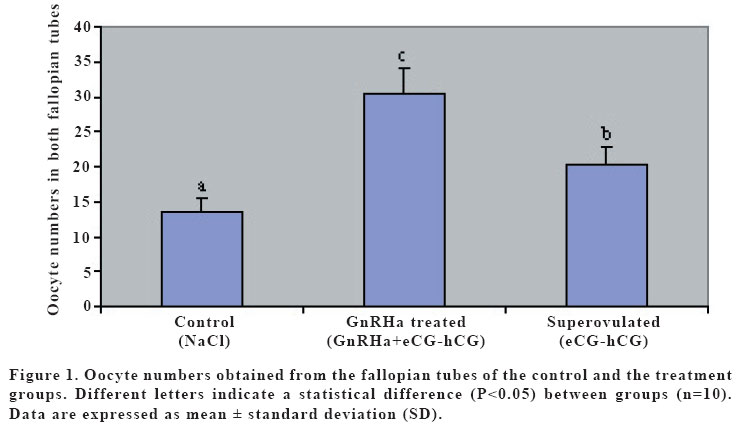
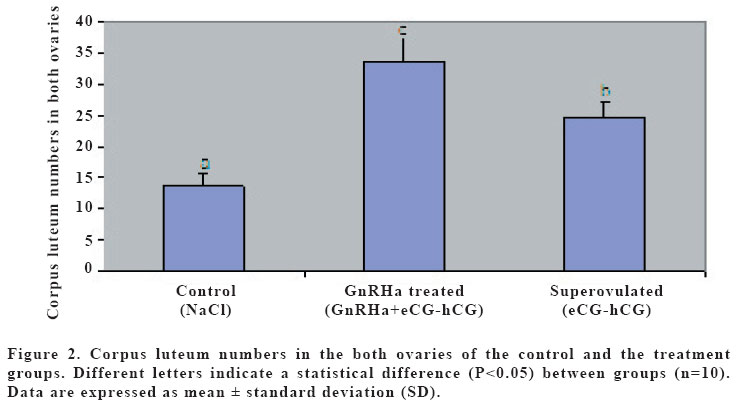
(eCG) and human chorionic gonadotropin (hCG). Keywords: GnRH agonist, Superovulation, Oocyte maturation, Mice. INTRODUCTION Gonadotropin-releasing hormone (GnRH) mediates the hypothalamic control of pituitary gonadotropin secretion and biosynthesis. However, recent studies have shown that, at least in the rat, GnRH and its agonist (GnRHa) could also elicit ovarian responses, both in vitro and in vivo (1). The effects of GnRH on the rat ovary include stimulation of oocyte maturation (2,3). However, direct gonadal effects of GnRH may interfere with the stimulatory effects of exogenous gonadotropin on follicular development, corpus luteum establishment, and oocyte maturation (4). It is well established that the follicular oocyte in mammals is arrested at the prophase of the first meiotic division and that the physiological stimulus for resumption of meiosis is provided by the preovulatory surge of luteinizing hormone (LH) (5). Native GnRH and its analogs may mimic LH action, thereby inducing resumption of meiosis; in that regard, in vitro exposure of isolated ovarian follicles to GnRH or its agonists resulted in oocyte maturation, via activation of specific GnRH receptors on granulosa cells (6,7,8). The aim of this study was to investigate the effect of a GnRH agonist on ovulation rate, and the number and maturation of oocytes in mice superovulated with eCG and hCG. MATERIALS AND METHODS Thirty female mice (BALB/C), aging 3 months and weighing between 25-30 g, were used. The mice were assigned to the three experimental groups of control, superovulated and GnRHa treated, each containing 10 animals, and fed with standard feed and water ad libitum. In order to maintain stable biological rhythms, 12 h of artificial light and 12 h of darkness were provided. All mice received humane care according to the criteria outlined in the “Guide for the Care and Use of Laboratory Animals”prepared by the National Academy of Sciences and published by the National Institutes of Health. Control mice were received only the intraperitoneal (i.p.) injection of 0.1 ml 0.9% NaCl. Superovulated mice were given an i.p. injection of 5 IU of equine chorionic gonadotropin (eCG; Folligon, Intervet, Istanbul, Turkey) with 0.1 ml 0.9% NaCl followed 48 hr later by 5 IU of human chorionic gonadotropin (hCG; Pregnyl, Organon, Istanbul, Turkey). The GnRHa treated mice were received an intramuscular (i.m.) injection of a 20 mg/kg GnRHa (Buserelin acetate:D-Ser-[TBU]6-des-Gly-NH210) LHRH ethylamide, obtained by Hoechst Japan (Tokyo, Japan) 24 hr before the superovulation with eCG-hCG. Thirteen hr after hCG administration, mice were sacrificed by cervical dislocation. The ovaries and the fallopian tubes were also harvested for counting the corpora lutea and the cumulus-enclosed oocytes, and put in a human tubal fluid (HTF)-medium (9). Corpora lutea on ovaries were counted under a stereo microscope (Nikon SMZ800, Japan). Then, the fallopian tubes were excised and the cumulus-enclosed oocytes were recovered from the tubes into HTF-medium containing 0.5 mg/ml hyaluronidase (Sigma Co., St Louis, MO). The oocytes were removed from the cumulus cells by gentle agitation using a narrow pipette and were placed in a fresh HTF-medium to count and determine the maturity of oocytes under an inverted microscope (Nikon T300, Japan). Oocyte maturity was based on the presence of polar body. The data were analyzed by using analysis of variance (ANOVA). Duncan test was used to test for differences among means for which ANOVA indicated a significant (p<0.05) F ratio. RESULTS Numbers of mature oocytes, and ovulation rate were higher (P<0.05) in GnRHa treated mice than controls and superovulated mice (Table 1). Although 88% of the oocytes were in the form of oocyte II (mature: with polar body) and 12% of the oocytes were in the form of oocyte I (immature: with germinal vesicle) in GnRHa treated mice, 60% of the oocytes were in the form of oocyte II and 40% of oocytes were in the form of oocyte I in superovulated and control mice. The numbers of the oocytes and the corpora lutea were higher (p<0.05) in GnRHa treated mice (Figures 1 and 2) than controls and superovulated mice. The oocytes obtained from all groups were in normal morphological appearance. DISCUSSION Pretreatment with GnRH prior to superovulation significantly increased the number of oocytes recovered, compared to superovulation without GnRH. Until fairly recently it was generally accepted that the pituitary, which stimulated by GnRH to synthesize and secrete gonadotrophins, is the only target for this hormone. However, later experiments have shown that GnRH and its GnRHa can also elicit ovarian cells (10,11). However, recent studies have shown that, at least in mice and rabbits, GnRH and its analogues also exert a direct effects of gonadal function, affecting or influencing oocyte mauration both in vivo and in vitro (3,12). It has been suggested that GnRH induces oocyte maturation via activation of specific GnRH receptors on granulosa cells (2,13). GnRHa induce meiotic maturation by mechanisms independent of ovarian prostaglandin (PG)E2 and PGF2α synthesis. These data support the hypothesis that GnRH or GnRHa may play a modulating role in the control of meiotic maturation at the ovarian level in rabbits (7). This is consistent with other data showing GnRHa stimulate in vitro synthesis of PGs in rat granulosa cells (11). Increasing concentration of PGs play an important role in oocyte maturation (14). GnRHa, unlike LH or hCG, did not influence cyclic AMP accumulation in rat granulosa cells (11) or in rabbit preovulatory follicles. GnRHa, therefore, would appear to stimulate PG accumulation by a mechanism that does not involve cyclic AMP. One cannot exclude the possibility, however, that the direct gonadal effects of GnRHa, including the stimulation of meiotic maturation and PG accumulation, may represent secondary pharmacological properties of this class of compounds (7). The increased number of mature oocytes in the GnRH-treated group in the present study also supported the idea that GnRH stimulated the resumption of meiosis in ovarian follicles. However, since we did not measure cyclic AMP concentration, we do not know whether it played a role in the GnRH-induced increase in oocyte number and maturation. In the present study, increased number of oocytes was associated with the normal morphological shape of these oocytes in GnRHa treated mice. This result indicated that the mature oocytes obtained from the GnRHa treated mice might be used in in vivo and in vitro fertilizations. Therefore, we concluded that an i.m. injection of the GnRHa 24 hr before the superovulation with eCG-hCG increases the oocyte number and the maturation. Increased number of mature oocyte will increase the chance of fertility. REFERENCES
Copyright 2005 - Medical Investigations Society The following images related to this document are available:Photo images[gm05001f2.jpg] [gm05001f1.jpg] [gm05001t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}