 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
European Journal of General Medicine, Vol. 2, No. 2, 2005, pp. 69-75 DOSE DEPENDENT EFFECTS OF CAFFEIC ACID PHENETHYL ESTER ON HEART RATE AND BLOOD PRESSURE IN RATS# Mustafa Iraz1, Ersin Fadıllıoğlu2, Seda Taşdemir1, Burhan Ateş3,Selim Erdoğan4 İnonu University, Faculty of Medicine, Departments of Pharmacology1 and Physiology2, Faculty of Art and Science, Departments of Chemistry3 and Analytical Chemistry4, Malatya, Turkey Correspondence: Mustafa Iraz, MD Inonu University, Faculty
of Medicine, Department of Pharmacology, 44280, Malatya, Turkey #This study was presented in “The Experimental Biology 2005 Annual Meeting and the XXXV International Congress of Physiological Sciences in San Diego, CA, 31 March- 6 April 2005” Code Number: gm05014 Aim: Caffeic acid phenethyl ester (CAPE) is one of the
major components of honeybee propolis and its structure is similar to flavonoids.
The molecular mechanisms of the effects of CAPE on various systems including
cardiovascular system have not been known well. The aim of the present study
was to investigate the short term dose dependent in vivo cardiovascular
effects including heart rate and blood pressure changes induced by CAPE in
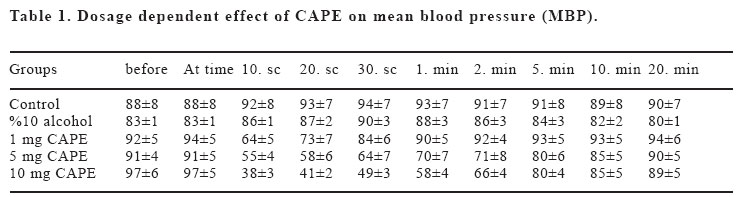
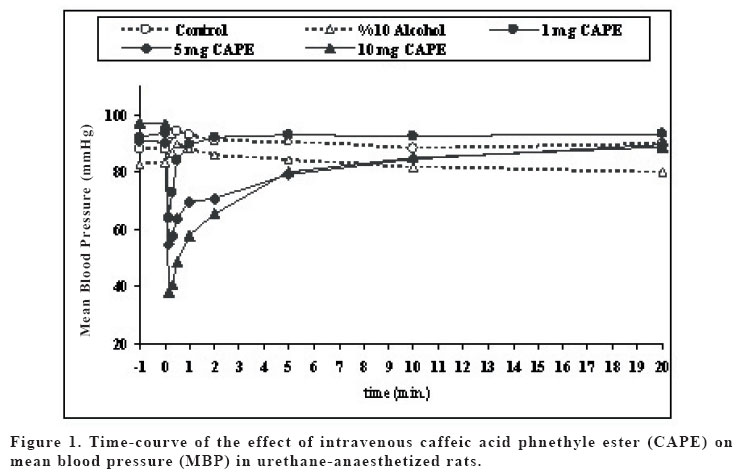
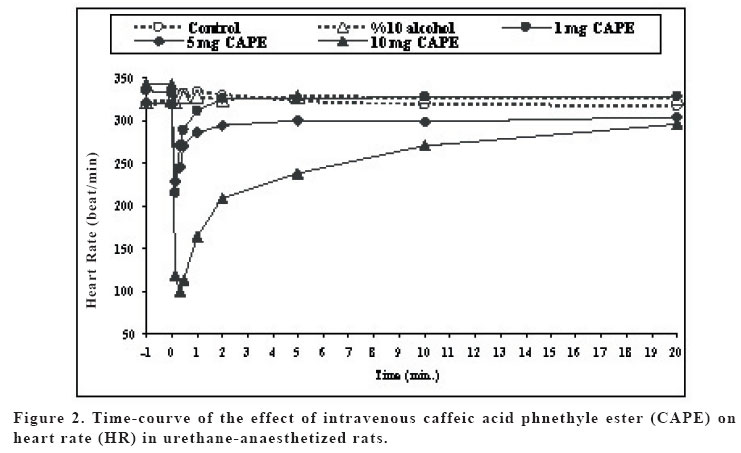
Sprague Dawley rats. Key words: Caffeic acid phenethyl ester (CAPE); bradycardia; hypotension; rat INTRODUCTION Propolis (bee glue) is known as a resinous dark-coloured material which is collected by honeybees from the buds of living plants mixed with bee wax and salivary secretions. Crude extracts of propolis contains amino acids, phenolic acids, phenolic acids esters, flavonoids, cinnamic acid, terpenes and caffeic acid, and its compositions alter resulting from variation in geographical and botanical origin (1). Propolis became a part of folk medicine and its biological effects, including anti-inflammatory, antiviral, antibacterial, immunostimulatory and carcinostatic activities, were evaluated in animal studies (2,3). The broad spectrum of activity of propolis was mainly attributed to the large number of flavonoids (4). Caffeic acid phenethyl ester (CAPE) is one of the major components of honeybee propolis and its structure is similar to flavonoids. CAPE has been demonstrated to have some biological and pharmacological properties, such as antioxidant (5,6), antiinflammatory (7,8), anticarcinogenic (9-11) and immunomodulatory (12) activities similar to flavonoids. The molecular mechanisms of the effects of CAPE on various systems including cardiovascular system have not been known well. The anti-inflammatory properties of CAPE have been attributed to suppression of prostaglandin (PG) and leukotriene synthesis (13). In recent years, Natarajan et al. (14) demonstrated that CAPE completely and specifically blocked the activation of nuclear transcription factor kappa B (NF-kB) and induced apoptosis; effects of inhibition of activation of NF-kB has been shown to account for the beneficial effect of this compound in a rat model of vascular injury (15). Moreover, CAPE has been shown to prevent ischemia reperfusion injury by means of a decreased NO level (16-18). Fadillioglu et al. (19) shown that CAPE prevented doxorubicin induced cardiotoxicity and restored the blood pressure changes due to doxorubicin cardiotoxiticy. In addition, Cicala et al. (20) demonstrated that CAPE inhibited contractile responses to phenylephrine or to KCl, and also inhibits the contractile response to phenylephrine obtained in a Ca+2-free medium. Thus, the aim of the present study was to investigate in vivo dose dependent cardiovascular effects including heart rate and blood pressure changes induced by CAPE in rats. MATERIAL AND METHODS In the experiment, 90 day old male Sprague Dawley rats weighing 250-300 g were used. Animals were housed two per cage in the animal quarter with an alternating 12 h light-dark cycle, air-conditioning room with 25oC temperatures and they were given standard rat pellet feed and water ad libitum. The experiments were performed in accordance with “Guide for the Care and Use of Laboratory Animals, DHEW Publication No. (NIH) 85–23, 1985”and approved by local ethical committee at Medical School of Inonu University. Rats were randomly divided into six groups (n:6) as follows: group 1 and group 2 were given serum physiologic (0.9% NaCl solution) and vehicle for CAPE (10% ethanol solution). Groups 3, 4, 5 and 6 were treated with 1, 5, 10 and 20 mg kg-1 with CAPE, respectively. CAPE applied in this study was synthesized according to the technique described by Grunberger et al. (21). 20 mg CAPE dosage caused death within seconds (sec), so that this group was discharged from the study. The rats were anaesthetized with i.p. urethane 1.2-1.4 g kg-1 and placed in the supine position with their head fixed to table coated with heating pad which kept body temperature constant at 37oC, and then tracheotomy was performed in order to allow the animals breathe spontaneously. The right jugular vein was cannulated with PE-50 polyethylene tube for intravenous (i.v.) administration of drugs. The left common carotid artery was cannulated with PE-50 tube. Blood pressure was measured in the common carotid artery with a BBT pressure transducer and ECG monitored (BIOPAC MP-100 data acquisition system, USA). The rats were left undisturbed until heart rate and blood pressure stabilized. Mean arterial blood pressure and heart rate were calculated from standard ECG records. The effects of serum physiologic, vehicle and different dosage of CAPE on mean arterial blood pressure were monitored during five min. prior to drug administration and after 60 min. following stabilization of blood pressure and heart rate (baseline period), drugs were administered as described above. Mean arterial blood pressure and heart rate were measured at following time schedule: one min. and just before drug administration; 10, 20 and 30 sec. and 1, 2, 5, 10, 30, 45 and 60 min. after drug administration. Statistical analyses were carried out using SPSS 10.0 statistical software (SPSS for Windows; Chicago, III., USA). Distribution of the groups was analyzed with one sample Kolmogrov-Smirnov test. All groups showed normal distribution, so that parametric statistical methods were used to analyze the data. One-way ANOVA test was performed for comparison. Post Hoc multiple comparisons were done with LSD. Repeated measurements of ANOVA were used to calculate interaction between interventions and than means of paired sample t-test (alterations in the same group) with Holm’s sequential Bonferroni procedure were applied for within group comparisons. p-values less than 0.05 were considered to be significant. RESULTS The mean blood pressure (MBP) values are shown in Figure 1. There were no significant differences between groups in MBP values before injections. 1 mg CAPE group had lower MBP than that of control group at 10. and 20. sec., and than that of vehicle group at 10. sec. (p<0.05). MBPs of 5 mg CAPE group were decreased in comparison with control and vehicle groups at 10., 20. and 30. sec. and 1. min; and control group at 2. min. (p<0.05). 10 mg CAPE group MBPs were lower than control and vehicle groups at 10., 20. and 30. sec. and 1.and 2. min (p<0.05). Also, 10 mg CAPE group MBPs were decreased in comparison with 1 mg and 5 mg CAPE groups at 10. and 20. sec. (p<0.05). MBPs of 5 and 10 mg CAPE groups were significantly lower than 1 mg CAPE group at 30. sec., 1. and 2. min. (p<0.05). The effects of CAPE on MBP were disappeared 5 min after injections (p>0.05). The heart rates (HR) are shown in Figure 2. There were no significant differences in HR between groups before injections and between control and vehicle groups during the experimental procedure. Control and vehicle HRs were significantly higher than all CAPE groups at 10. sec. (p<0.05). HRs were decreased in 5 and 10 mg CAPE groups in comparison with control and vehicle groups at 10. sec. (p<0.05). 10 mg CAPE administration caused significant decrease in HR compared to control and 10% alcohol groups at 30. sec., 1., 2. and 5. min. (p<0.05); and 1 mg CAPE group at 10., 20. and 30. sec., 1., 2. and 5. min. (p<0.05); and 5 mg CAPE group at 10., 20. and 30. sec., 1. and 2. min. (p<0.05). The effects of CAPE on HR were disappeared 10 min after injections (p>0.05). HR in 5 mg CAPE group was significantly decreased from prior to injection to 10. and 20. sec. (p<0.05), after that time HR level was lasted non-significantly lower than baseline level. HR in 10 mg CAPE group was significantly decreased in 10., 20., 30. seconds 1. and 2. min (p<0.05), compared to pre-injection level. 1 mg CAPE administration caused non-significant decrease at 10. 20. sec. compared to HR prior to injection. There was no significant change in HR in control and vehicle groups during experimental procedure. MBP in 10 mg CAPE group was significantly decreased in 10., 20. and 30.seconds 1., 2. and 5. mins. compared to beginning baseline level (p<0.05). MBP in 5 mg CAPE group was significantly lower at 10. and 20. sec. after drug administration than beginning baseline level (p<0.05). 30 sec. after CAPE administration and afterwards, MBP was non-significantly lower than baseline level. MBP in 1 mg CAPE group was significantly decreased at 10 sec. after the drug administration in comparison with beginning baseline level of MBP (p<0.05). Then, MBP returned to baseline level in 1 mg CAPE group. DISCUSSION The present study demonstrated that i.v. CAPE treatment resulted in changes in both heart rate and blood pressure. The hypotensive effect of CAPE in this study was in agreement with previous studies. It was shown that CAPE ameliorated the hypertensive effects of cardiotoxic agent(19). We also measured low heart rate during CAPE treatment in rats. CAPE caused hypotensive effects with low HR in our study. Several studies had shown the vascular effect of CAPE. It was demonstrated that in vitro CAPE caused both concentration and endothelium dependent relaxation of rat thoracic aortic rings precontracted with phenylephrine or with KCI, and reduced the contractile response to phenylephrine or to KCI by Cicala et al.(20). Furthermore, CAPE reduced phenylephrine-induced contraction in a Ca2+–free medium and abolishes phenylephrine- and KCI- induced intracellular Ca2+ increase. Also, they demonstrated the relaxant effect of low concentrations of CAPE and this was abolished when tissue was incubated with L-NAME, or derived of endothelium, while the effect of high concentrations was unaltered. Their results suggested that CAPE acts via an NO-dependent and an NO-independent mechanisms. In addition, their study showed that on endothelium-denuded vessels, relaxation induced by CAPE was not affected by inhibition of adenylate cyclase, ruling out any involvement of cAMP (20). Also, previous studies demonstrated that ethanolic extract of propolis which was include CAPE and the others components, inhibited the contraction of trachea smooth muscle induced by histamine, capsaicin, KCI and carbacol, a muscarinic agonist, in vitro (22). The observed effect of propolis extracts was associated with the control of calcium mobilization, because KCl and histamine stimulated the opening of Ca2+ channels. Indeed, in recent years, the effects of CAPE on ion current have been focused. Our results showed that CAPE resulted in decrease in blood pressure. However the underlying mechanism has not been clear yet. NO or ion channel control is placed in the mechanism of hemodynamic changes due to CAPE. Ion channels are involved in neuronal control mechanism which is fast and short acting. But, vascular control mechanisms are mostly due to local factors. Our results showed that effects of CAPE were to be over within seconds or minute. So that, we thought that CAPE might affect the MBP mostly by neuronal way. Parker et al. (23) suggested that high dosage of CAPE reduced interleukin-1β induced interleukin-6 (IL-6) release. The synthesis and release of IL-6 and PGE2 from glial cells are important in the central nervous system (CNS) inflammatory response to infections and injury. The present data demonstrated that effect of i.v. CAPE with high dosage such as 20 mg kg-1 caused death. We thought that the reason of the death might be hemodynamic shock due to low blood pressure and HR. Oral CAPE administrations did not caused death (18). It might be metabolized during i.p. and oral ways, however i.v. administrated CAPE reaches directly to the target tissues such as heart, vascular beds and neuronal systems. Fadillioglu et al. (24) demonstrated that CAPE prevented doxorubicin induced cardiotoxicity. Doxorubicin caused high blood pressure and high HR and CAPE returned the hemodynamic changes to the baseline level. Also, it is shown that CAPE inhibited NO synthesis gene and enzyme activity (25) and returned the high NO production during DXR-administration(24). Nitric oxide related vascular effect of CAPE may not be rule out. Because NO is produced as a neurotransmitter other than an inflammatory way. The control of heart rate depends on afferent inputs from visceral afferents. It has been shown that NO can act on peripheral afferent excitability with effects of the nucleus tractus solitarii (NTS) which are the brainstem termination site for baroreceptor afferents. It was demonstrated that NO modulates release of neurotransmitters, such as GABA and glutamate (26). The control of cardiovascular system through NTS may be carried out with modulation of these neurotransmitters. Normally, NO in nervous tissue is produce by two enzymes, neuronal NOS (nNOS) and eNOS (26). So that, effect of CAPE on iNOS activity may not deny the role of NO on heart rate and blood pressure through control of NTS during CAPE administration. CAPE has blocking effects on iNOS activity and NF-kB to prevent inflammatory answer of the tissue to the stress. However, this decreasing effect on NO production via anti-inflammatory action may not put away the cardiovascular regulation of CAPE through NO. CAPE may act via CO and heme oxygenase (HO) system. Carbon monoxide induces cGMP system in cellular mechanism especially in smooth muscle cells in vasculature. Carbon monoxide derived from HO contributes to modulate important physiological processes within the cardiovascular system which includes the regulation of vessel tone (27). Carbon monoxide acts as a neurotransmitter and may have a role in the regulation of vascular tone. CO has a function as an endogenous modulator of the NO-cGMP signaling system in the brain. NO is able to cause high HO-1 expression and activity in tissue. Motterlini et al. (27) showed the scientific evidence which supports the role for HO in suppression of acute hypertension. Johnson et al. (28) showed that administration of zinc protoporphyrin IX, a potent heme oxygenase inhibitor, caused an increase in arterial pressure in rats. CAPE is known to be specific inhibitors of nuclear transcription factor NF-kB and cyclooxygenase activity. CAPE also inhibits lipid peroxidation and has a capacity to prevent the oxidation of important intracellular components. CAPE has a preferential affinity toward selective cysteine residues of targeted proteins that finely control the transcription of inducible genes. Scapagnini et al. (29) demonstrated that CAPE caused marked increase HO activity in astrocytes as a novel and potent HO-1 inducer. Effect of CAPE on regulation of blood pressure and heart rate may be due to effect on HO and CO systems in brainstem. The increase in tissue HO-1 expression and activity correlated with increased CO production from aorta and urinary bilirubin excretion, indicating a functional link between tissues HO-1, its active function as a heme degrading protein, and its physiological role in blood pressure regulation (27). HO-1 is an inducible protein which may be protecting the brain from oxidative damage and its expression was demonstrated to be critical in modulating the response of neurons to various kinds of stress (30). The hemodynamic effect of CAPE is similar to the symptoms of mad honey intoxication which includes bradycardia and hypotension. Grayanotoxin compound of mad honey responsible for the intoxication (31,32). However, it has not been known CAPE contents of mad honey and responsibility for its intoxication. CAPE by i.v. route induces immediate and dose dependent hypotension and low HR. To be the best of our knowledge, this is the first time that such hypotensive and bradycardic effects of CAPE have been reported in rats in vivo. CAPE may affect the NTS via HO-1 and CO system as well as NO system. Although the exact mechanisms remain to be clarified, CAPE could be an effective cardiovascular therapeutic agent. Further investigations are needed regarding CAPE, NO and other vascular factors to clarify the exact mechanisms of cardiovascular effects. Also, effects of CAPE on ion channels, especially on excitability cells which control the cardiovascular autonomic system, should be investigated. REFERENCES
Copyright 2005 - Medical Investigations Society |

{kind=link}
{kind=link}
{kind=link}
{kind=link}