 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
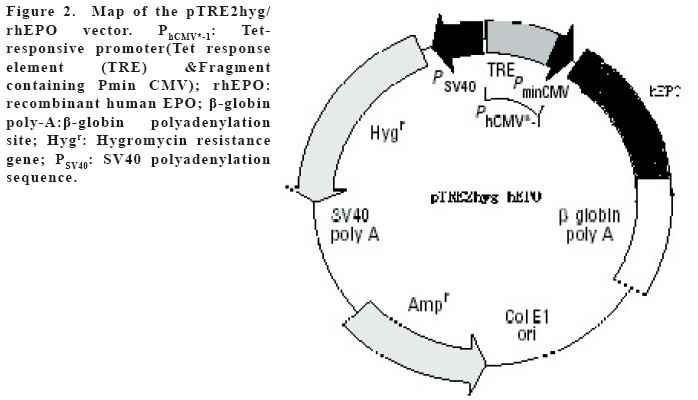
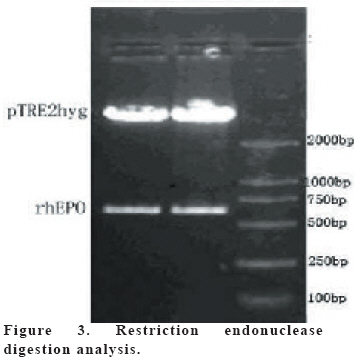
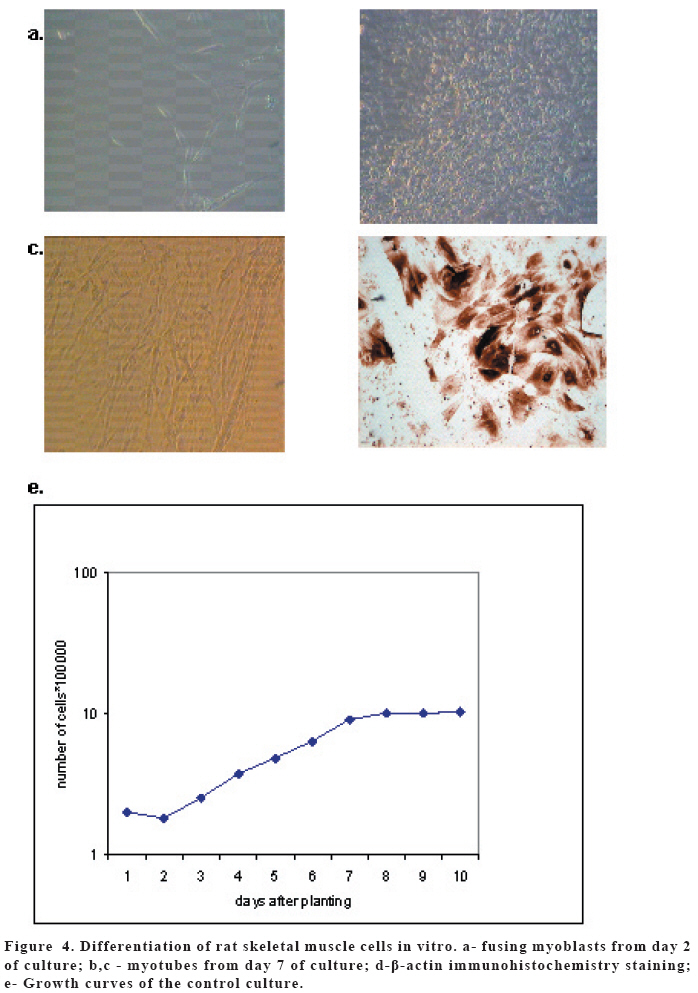
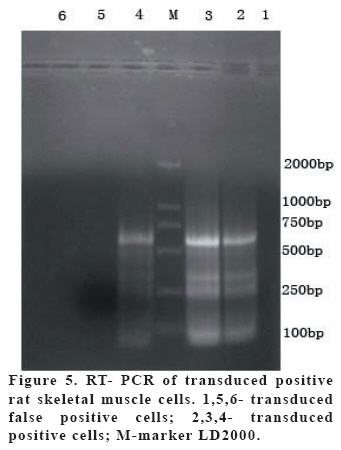
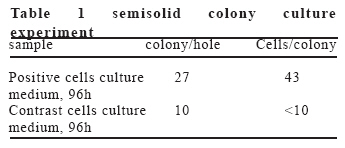
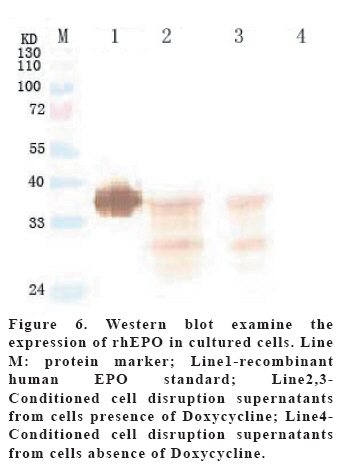
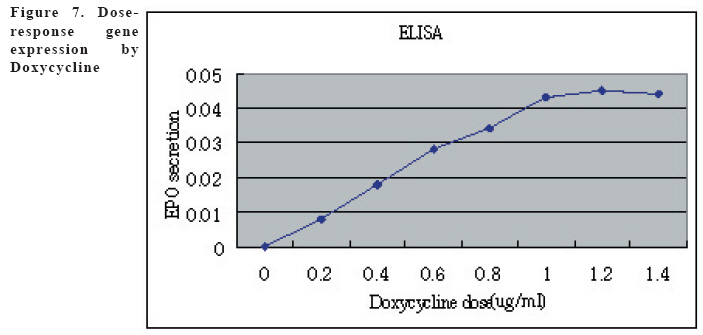
European Journal of General Medicine, Vol. 3, No. 3, 2006, pp. 108-115 REGULATION OF ERYTHROPOIETIN EXPRESSION BY DOXYCYCLINE IN RAT PRIMARY SKELETAL MUSCLE CELLS FOLLOWING TRANSDUCTION BY TETRACYCLINE GENE EXPRESSION SYSTEM Dai Yong1, Xu Zhuojia, Li Tiyuan 2nd Affiliated Hospital of Medical College of Jinan University, Shenzhen People’s Hospital, China Correspondence: Dai Yong 2nd Affiliated Hospital of Medical College of Jinan University, Shenzhen People’s Hospital, Shenzhen 518020. China Phone: 86 13802201510 Fax: 86 075525626750 E-mail: daiyong22@yahoo.com.cn Code Number: gm06023 Aim: Future prospects for gene therapy of renal anemia involve expression of the EPO transgene. Control of gene expression is important to gene therapy for purposes of both dosing and safety. We tested the effects of rhEPO gene expression under the tetracycline gene expression system’s regulating and controlling. Key words: EPO, Dox, Gene therapy, Skeletal Muscle cells . INTRODUCTION Erythropoietin (EPO) is a 30-kd glycoprotein hormone that is the regulator of red blood cell production and maintenance in mammals. (1,2) In human adults, the erythropcyte-generating hormone, is produced mainly by the kidney peritubular cells (3). Anaemia is a major complication of chronic renal failure (CRF), which results from a destruction of EPO-secreting cells. The administration of recombinant EPO is now widely used for long-term treatment of anemia associated with chronic renal failure, cancer chemotherapy, and HIV infections. Currently, about 300,000 CFR patients throughout the world are treated with recombinant human EPO. (4) However, this treatment requires repeated administration of recombinant protein, which is both inconvenient and expensive (5). Delivery of this hormone by gene therapy rather than by repeated injections may provide clinical and economic benefits and would serve as a model for the expression of other therapeutic proteins. Gene therapy offers the potential for increased efficacy and more cost-effective production and administration of a variety of genes coding for therapeutic proteins (6-12). Gene delivery to skeletal muscle (13, 14), vascular smooth muscle (15), and liver (16) can result in systemic levels of therapeutic proteins. Skeletal muscle is a useful target to evaluate this approach because of its large mass, vascularity, and accessibility. Since muscle fibers are nondividing, effective gene delivery could potentially result in long-lived protein production. Posttranslational modification of proteins in muscle cells allows these proteins to be secreted with full potency and bioavailability (17). Indeed, genetically modified myoblasts are capable of long-term delivery of growth hormone (13, 14), coagulation factor IX (18), erythropoietin (19), and β-glucuronidase (20). Although these studies are encouraging, there has been no convincing demonstration that this approach will be clinically useful or practical. More effective methods for gene delivery would be desirable to achieve sustained high-level expression of therapeutic gene products (21). Vectors of the tetracycline-dependent transcriptional regulatory system(22-25) were generated in which the tetracycline transactivator or the reverse tetracycline transactivator are controlled by the Tet-responsive PhCMV-1 promoter. Dominant selectable cassettes were introduced into each plasmid, allowing for selection following gene transfer into cells by incorporating hygromycin and G418 into the medium. To model an essential gene under tetracycline regulation, the hygromycin resistance gene (hph) was placed under the control of seven copies of the TetR binding site (tetO) in a plasmid vector and co-transfected into cells together with one of the two transactivator plasmids. Since the hph gene is essential to cells in the presence of hygromycin, resistance to hygromycin was used as a marker of hph reporter gene expression. Transformants were identified in which the expression of tTA conferred hygromycin resistance by activating expression of the tetO-hph reporter gene(26). MATERIAL AND METHODS Vector construction. The rhEPO cDNA was cloned by RT-PCR from CHO cells (Chinese Hamster Ovary cells), which contain rhEPO gene (YONGMING limited company donate). Total RNA was prepared from CHO cells using Trizol reagent (Sangon, Shanghai, China) according to the manufacturer’s instruction. First strand cDNA was prepared with oligo(dT) primer and Superscript reverse transcriptase (Sangon, Shanghai,China)by the provided protocol. The rhEPO cDNA was amplified PCR using Taq polymerase and standard reaction conditions. The sense and antisense primers were 5’CG GGA TCC GCC ACC ATG GGG GTG CAC GAA T3’ 5’CG ATC GAT TCA TCT GTC CCC TGT CCT3’. PCR products were cloned into the multiple cloning site (MCS) of pTRE2hyg (B&D, USA), which digested with BamH I and Cla I and sequenced. Rat Skeletal Muscle Cells culture. Skeletal muscle cell samples of 10 new-born SD rat derived from tissue pieces and digested with trypsin, cultured with two kinds of culture methods, the cells were placed in fusion medium (DMEM/10% Fetal calf serum/1% penicillin/streptomycin). The muscle cells were tested by β-actin immunohistochemistry stains. And culture cells with medium, which includes different degrees of G418 and homomycin B. Transfections. Rat skeletal muscle cells (106 cells in a 10-cm tissue culture dish) were co- transduced with each 0.5μg of pTRE2hygrhEpo and Tet-On plasmids using the LIPOFECTAMINE 2000 reagent (Invitrogen). Approximately 6 hours later, the transduced cells were placed in fusion medium (DMEM/10% Fetal calf serum/1% penicillin/streptomycin) and the cells were allowed to fuse into myotubes overnight. Two days later, added G418 (800 μg/ml) and homomycin B(200 μg/ml) into fusion medium for direct selection of stable transformants. About 10-15days, selected transduced positive cells and expanded the positive cells. Added Dox (1μg/ml, SIGMA,USA) in the culture medium. EPO Function Measurement. Positive cells and control cells culture medium were harvested after 96 h and harvest these cells to measure rhEPO gene expression and biological activation of EPO by RT-PCR (same as Vector constructs part) and semisolid colony culture experiment. Western Blot Analysis. Culture supernatants from transduced positive rat skeletal muscle cells (5 pfu per cell) were concentrated 20-fold using a Centricon-10 filter (Amicon). An aliquot of the concentrated supernatant was denatured and subjected to digestion with 0.5 unit of N-Glycanase (Genzyme) for 18 h at 37°C according to the manufacturer's suggested protocol. Untreated and N-Glycanase-treated myotube culture supernatants were fractionated by SDS/PAGE, transferred to nitrocellulose, and subjected to Western blot analysis using a commercially available polyclonal anti-hEPO antibody (R & D Systems). Regulation of gene expression. Added different concention Dox (0-1.4μg/ml ) in positive cells and cell medium were harvested after 96 h . The expression levels of EPO were measured by ELISA(R&D,USA) according to the manufacturer’s instruction.. RESULTS Construction and characterization of the pTRE2hyg/rhEPO. rhEPO coding sequences were obtained by RT-PCR(Figure 1) and inserted in pTRE2hyg vector. Expression was controlled by Tet-responsive PhCMV-1 promoter. PhCMV-1 is silent in the absence of blinding of rTetR to the tetO sequences. PTRE2hyg/rhEPO also contains the hygromycin resistance gene for direct selection of stable transformants (Figure 2,3). Rat Skeletal Muscle Cells culture. We showed two kinds of culture methods could harvest adequate cells for gene therapy research use. Cell differentiated was verified by revealing β-action expression in myotubles. The cell proliferation arrived climax during 7 day after transfer of culture (Figure 4). EPO Measurement. Expression of the pTRE2hyg/rhEPO vectors was analyzed in transduced rat skeletal muscle cells by RT-PCR(Figure 5). Total RNAs were prepared in the presence of Dox, either from undifferentiated myoblasts or after differentiation into myoblasts. The biology activity EPO protein was measured by semisolid colony culture experiment shown that the secreted EPO protein can stimulus the cellula medullares proliferation (Table 1) . Western Blot Analysis. Conditioned cell disruption supernatants were collected, analyzed by Western blot. As shown in Figure 6, EPO mediated the production of immunologically cross-reactive material that comigrated with standard rhEPO. The presence of two talin bands was detected. The upper one (35 kDa) is for a native form of talin, while the lower one (30 kDa) was formed as the result of talin cleavage. The bands were present at almost the same level in homogenates from all the stages of the control and Dox-treated cultures. EPO was not detected in the supernatant of non-transduced cells. A dose-response gene expression. With the aim to document the cells specificity and the reversibility of Dox-induced gene expression, EPO secretion was measured at regular intervals during the skeletal muscle cells cotransduced with pTRE2hyg/rhEPO and Tet-On vectors. When doxycycline was added, EPO secretion increased, researching levels equivalent to that of cells permanently maintained in the presence of the drug (1.4μg/ml). These results indicated that EPO secretion levels could be regulated by Dox in cultures of transduced rat skeletal muscle cells (Figure 7). DISCUSSION Anemia of chronic renal failure CFR, which results from a destruction of EPO-secreting cells, is one of the more common blood disorders and can lead to a series of health problems. Currently, CRF patients throughout the world are treated with rhEPO. Erythropoietin protein therapy is the treatment of choice for anemia of renal failure. One strategy is that the rhEPO is injected directly. However, this treatment requires repeated administration of recombinant protein, which is both inconvenient and expensive. An actively pursued treatment strategy is to reactivate the EPO expression of the transgenic cells themselves with the regulation of drugs. Delivery of EPO by gene therapy rather than by repeated injections may provide clinical and economic benefits. The tetracycline-regulatable is extremely attractive simply because the drug used to induce transcription is a well-known antibiotic, essentially devoid of major secondary effects. Dox has been extensively used in patients and its pharmacological and toxicological characteristics are known. This system is fit to gene therapy purposes, and has been used widespread. In this study, hEPO cloned from CHO cells, was successfully constructed the vector under a tetracycline-dependent expression system, which was cotransduced successfully with Tet-On vectors into skeletal muscle cells. All positive cells displayed tightly regulated EPO synthesis through the analysis of induction kinetics, which showed the peak of EPO production occurred at 7 D after Dox induction, keeping on the level after 8 D, and EPO secretion increased, researching levels equivalent to that of cells permanently maintained in the presence of the drug (1.0μg/ml). Controlling gene expression is a desired goal of gene therapy, which can potentially monitor the therapeutic effect of the transgene. In this study, rhEPO secretion in the skeletal muscle cells can be effectively controlled, depending on the Dox the dose and the effected time. In conclusion, based on the experiments we have carried out, we are making efforts to establish an artificial skeletal muscle cells in response to Dox stimulation under the control. Meanwhile, the rhEPO in skeletal muscle cell dose-depended regulatory system was applied for the clinical trial, as the model for a new, practical and effective system of gene therapy. Abbreviation: Dox, doxycycline; EPO, erythropoietin; CHO cells, Chinese Hamster Ovary cells; tTA, tetracycline transactivator; hph, hygromycin resistance gene; rtTA, reverse tetracycline transactivator, tetO, TetR binding site; Pmin CMV , minimal CMV promoter REFERENCES
Copyright 2006 - Medical Investigations Society The following images related to this document are available:Photo images[gm06023f7.jpg] [gm06023t1.jpg] [gm06023f6.jpg] [gm06023f4.jpg] [gm06023f3.jpg] [gm06023f1.jpg] [gm06023f5.jpg] [gm06023f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}