 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
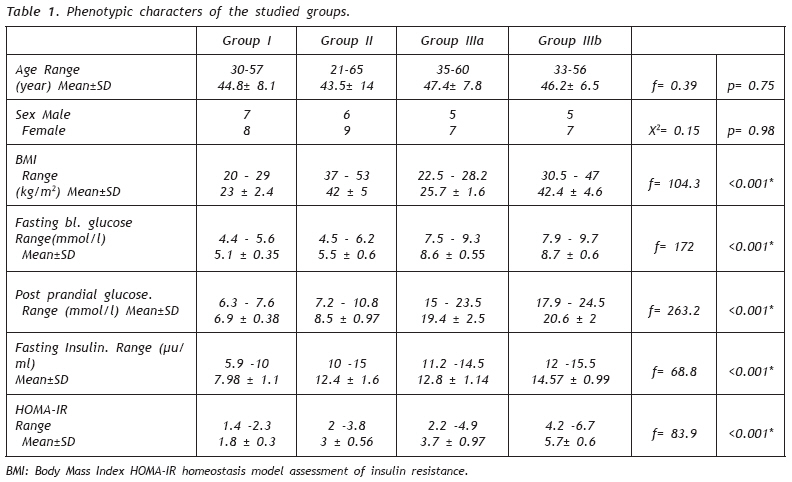
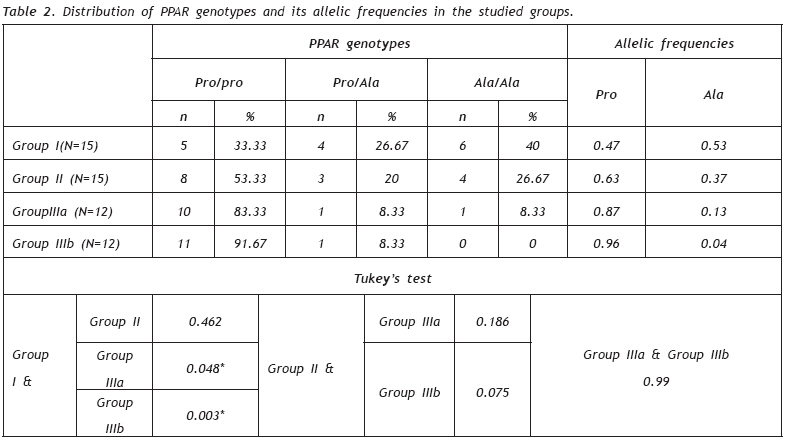
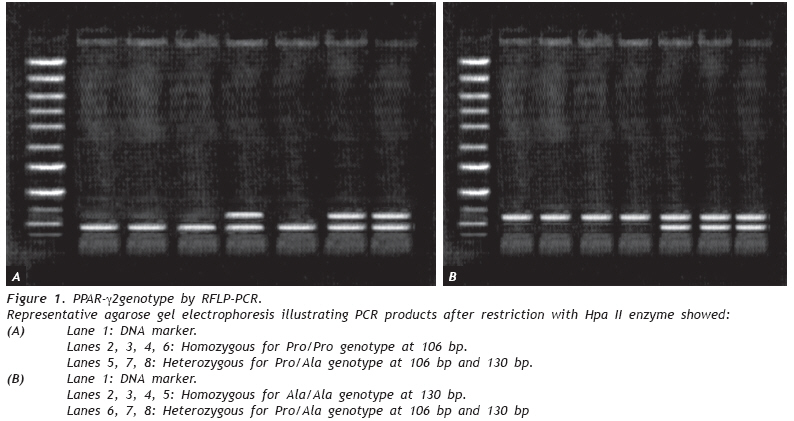
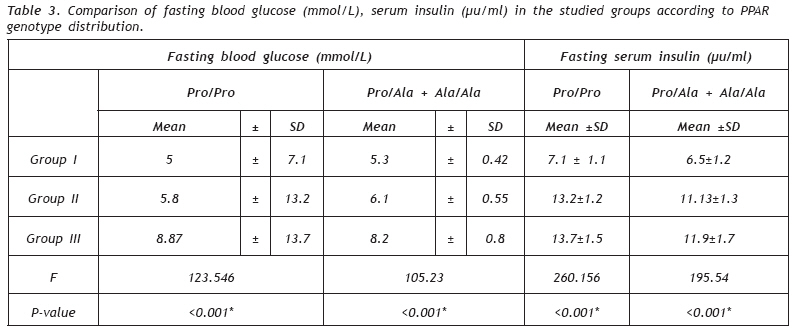
European Journal of General Medicine, Vol. 6, No. 2, 2009, pp. 78-86 Role of Peroxisome Proliferator- Activated Receptor gamma2 (PPAR-γg2) Gene Polymorphism in Type 2 Diabetes Mellitus Salwa Sokkar1 , Jehan A. El-Sharnouby1 , Amal Helmy1 , Amal El-Bendary1 , Laila Sayed Ahmad2 , Kamal Okasha2 1 Tanta University, Faculty of
Medicine,Department of Clinical
Pathology Code Number: gm09019 INTRODUCTION Diabetes mellitus is a group of metabolic disorders characterized by hyperglycemia resulting from defect in insulin secretion, insulin action or both (1). The pathogenesis of type 2 diabetes is multifaceted involving both lifestyle and genetic component. Insulin resistance in peripheral tissues is a high-risk factor and almost invariably precedes the development of overt type 2 diabetes (2). Insulin resistance is associated with obesity especially when centrally distributed. The connection between increased adiposity and insulin resistance is still poorly understood, although recent evidence has suggested that adipose tissue-released cytokines like adiponectin, resistin, leptin and Tumour Necrosis Factor-Alpha (TNF Alpha) may be contributory factors (3) that modulate insulin secretion and insulin action (4). Peroxisome proliferator-activated receptors (PPARs) are ligand-activated transcription factors belonging to the nuclear hormone receptor superfamily (5). There are three isoforms, encoded by separate genes: PPAR-[gamma], PPAR-[alpha], and PPAR-[beta]/ [delta] ,which share 60% to 80% homology in their ligand- and DNA-binding domains, and exhibits distinct patterns of expression and overlapping and distinct biological activities (6). There are three isoforms of PPARg have been identified by alternative promoter usage and splicing of PPAR gene, PPARg1, PPARg2, and PPARg3. Proteins produced from PPARg2 contain an additional NH2terminal region, composed of 30 amino acids. PPARg2 expression is primarily restricted to adipose tissue (7). PPARg plays a critical role in glucose homeostasis and is the molecular target of a class of insulin-sensitizing drugs called thiazolidinediones (TZDs), which are PPARg ligands and are widely used for treatment of type 2 diabetes (5). PPARg ligands also upregulate expression of adipocyte glycerol kinase, thereby stimulating glycerol incorporation into triglyceride, even under conditions which would normally favour lipolysis, and resulting in improved insulin action in target tissues such as liver and muscle (8). PPARg ligands are also shown to augment glucose disposal in peripheral tissues by increasing expression of the glucose transporter genes glucose transporter 1 (GLUT1) and GLUT4. They profoundly alter gene expression in adipose tissue. Expression of resistin and tumor necrosis factor [alpha] (TNFa), which both induce insulin resistance, are reduced by PPAR g ligands (9). In addition, expression and secretion of adiponectin, a hormone exclusively produced by the adipocyte and increases insulin senstivity, is increased in the presence of PPAR g agonists. These data suggest that the adipose tissue is the primary target of PPAR g ligands, resulting in an improvement in insulin sensitivity in liver and muscle (10). The human peroxisome proliferator activated receptor-[gamma] (hPPAR[gamma]) hPPAR g gene is located on chromosome 3 at position 3p25 (11). hPPAR genes characterized to date reveal a common organization of the translated region in six coding exons: one exon for the N-terminal A/B domain, two exons for the DNA-binding domain (DBD, one exon for the hinge region, and two exons for the ligand-binding domain (LBD) (7). PPAR g 2 is encoded in seven exons, the common six exons and specific PPAR-g2 exon (exon B) which encode additional N-terminal amino acids specific of PPARg2 (12). The human PPAR protein is formed of four domains. The most important of them are DNA binding domain (DBD) and ligand binding domain (LBD). The DBD can regulate transcription of target genes by forming heterodimers with the retinoid X receptor (RXR) which is a common DNA binding partner to many nuclear receptors of the steroid/thyroid receptor superfamily (13), and binding to specific PPAR response elements (PPREs) in the promoter region of target genes (5). The most important character of PPARs protein lies in their particular ligand binding properties, making them attractive therapeutic targets. The LBD is unusually large, allowing PPARs to interact with a broad range of structurally distinct natural ligands as unsaturated fatty acids and synthetic ligands as thiazolidinediones which are a class of antidiabetic drugs (14). Pro12Ala polymorphism is a frequent cytosine to guanine single nucleotide polymorphism in PPARg2 exon B (CCA-to-GCA missense mutation). This variation results in a Proline to Alanine substitution at the codon 12, which has been found to modulate the transcriptional activity of the gene (15). So, the aim of this work to investigate the possible role of PPAR-g2 gene polymorphism as a genetic risk factor for type 2 diabetes mellitus and also its possible role as a link between obesity and type 2 diabetes mellitus. MATERIAL AND METHODS This study included 54 subjects. They were classified into: Group I: Included 15 apparently healthy lean individuals (7 males and 8 females) as control group. Their age ranged from 30 to 57 years, Group II: Included 15 obese non diabetic individuals (6 males and 9 females). Their age ranged from 21 to 65 years and Group III: Included 24 patients with type 2 diabetes mellitus with no diabetic complications (10 males and 14 females). Their age ranged from 33 to 60 years .They were selected from outpatient clinic of Tanta University Hospital. This group divided into: Group IIIa: Diabetic non obese patients (12 patients) and Group IIIb: Diabetic obese patients (12 patients). All subjects in this study were subjected to full history taking especially the age of onset of diabetes mellitus in group III, duration of the disease and regimen of the treatment, full clinical examination with measuring the body weight and length for estimation of BMI which is calculated as body weight (BW, kg) divided by body height (BH, m2). Obesity is defined as a (BMI) of ≥ 30 kg/m2 and overweight is defined as (BMI ≥ 25 kg/m2) (16). Peripheral blood samples were collected from fasting patients and controls. Each sample was divided into 2 mL in blank tube for separation of serum for estimation of fasting glucose level and serum insulin hormone level, and 3 mL in sterile vacuum tubes containing EDTA for DNA extraction and studying of PPAR-g2 gene polymorphism. 2-h post prandial blood samples were also collected for glucose estimation. Fasting serum insulin level by ELISA using Diagnostic systems Laboratories (DSL), HOMA insulin resistance: The estimation of insulin resistance by homeostasis model assessment (HOMA-IR) was calculated with the formula: Fasting serum insulin (µIU/ml) X fasting glucose level (mmol/L)/22.5 (17). Normally in men → 1.66 ± 1.1 and in women → 1.57±0.8. -Estimation of PPAR-g2 gene polymorphism by restricted fragment length polymorphism-polymerase chain reaction (PCR) (18): DNA was extracted from blood samples by using V-Gene DNA isolation Kit, supplied by Biotechnology, according to the manufacturers’ instructions. The paired primers (Operon) for PPARg2 gene were: Forward primer 5`-CAAGCCCAGTCCTTTCTGTG-3` Reverse primer 5`-AGTGAAGGAATCGCTTTCCG-3` All reactions were performed in a total volume of 50µl containing 10 mmol/L Tris HCl (PH 8.8), 50 mmol/L KCl, 1.5 mmol/L MgCl2, 0.2 mmol of each dNTP, 50 pmol of each of the two primers, unit of Taq DNA polymerase (Stratagene), 200 µg of genomic DNA was added last to PCR. The amplification program was started as follow: Initial precycling denaturation by holding at 94°C for 3 minutes, denaturation for 40 cycles at 94°C for 30 seconds followed by annealing at 53°C for 30 seconds, extention at 72°C for 1 minute and a final extention period at 72°C for 9 minutes. Restriction fragment length polymorphism (RFLP) was detected after digestion over night with 2 U of the restriction enzyme Hpa II (New England Biolabs, Beverly, Mass., USA) which cuts the mutant allele at a site introduced by the reverse primer. Samples were applied to 3% agarose gel and subjected to electrophoresis for about about half an hour and visualized on a UV transilluminator. Statistical Analysis Statistical presentation and analysis of the present study was conducted using the mean, standard deviation, student t-test analysis of variance [ANOVA] test, Hardy-Weinberg proportions and chi-square test. Data were analyzed using SPSS software, version 11. Significance was set statistically at P-value less than 0.05. RESULTS Phenotypic characters of the studied groups illustrated in Table 1 including; age, sex, BMI, fasting blood glucose, postprandial blood glucose, fasting serum insulin and HOMA-IR. The distribution of PRAR genotype and it’s allelic frequency (Table 2, Figure 1). In group I. 5 individuals (33.33%) carrying Pro/Pro genotype, 4 individuals (26.6%) carrying Pro/Ala genotype and 6 (40%) carrying Ala/Ala genotype Allelic frequencies are 0.47 for Pro allele and 0.53 for Ala allele in this group. In group II 8 individuals (53.33%) carrying Pro/Pro genotype, 3 individuals (20%) carrying Pro/Ala genotype and 4 (26.67%) carrying Ala/Ala genotype and allelic frequencies are 0.63 for Pro allele and 0.37 for Ala allele in this group. In group IIIa 10 individuals (83.33%) carrying Pro/ Pro genotype, 1 individuals (8.33%) carrying Pro/Ala genotype and 1 (8.33%) carrying Ala/Ala genotype and allelic frequencies are 0.87 for Pro allele and 0.13 for Ala allele in this group. In group IIIb 11 individuals (91.67%) carrying Pro/Pro genotype, 1 individuals (8.33%) carrying Pro/Ala genotype and non carrying Ala/Ala genotype and allelic frequencies are 0.96 for Pro allele and 0.04 for Ala allele in this group. Significant increase of Pro allele was found in group IIIa, group IIIb when compared to group I (P=0.048 and 0.003, respectively). However, there was no difference between group II and group I as regard the allelic frequencies (P=0.462). While group IIIb had higher Pro allele frequency (p= 0.003) than in group IIIa (p= 0.048). The fasting blood glucose levels had no difference between Pro/Pro carriers and Pro/Ala plus Ala/Ala carriers in groups I, II and III (p= 0.17, 0.31 and 0.126; respectively) (Table 3). Also, the mean fasting serum insulin level was significantly increased in Pro/Pro carriers when compared to that of Pro/Ala plus Ala/Ala carriers (p= 0.39, 0.01 and 0.026; respectively in groups I, II and III). HOMA-IR was significantly increased in Pro/Pro carriers in group II and group III when compared to that of Pro/Ala plus Ala/Ala carriers (p= 0.004 and 0.023; respectively) (Table 4). As regard the body mass index (BMI) there was no difference between Pro/Pro carriers and Pro/Ala plus Ala/Ala carriers (p= 0.58, 0.4 and 0.14; respectively in groups I, II and III). DISCUSSION The current study was designed to find out the relation between Pro 12 Ala polymorphism of PPARγ2 gene with type 2 diabetes mellitus and the possible role of this gene polymorphism as a link between obesity and type 2 diabetes mellitus by using polymerase chain reaction. Significant increased in Pro allele was found in diabetic non obese patients& diabetic non obese patients when compared to control group (p= 0.048 and 0.003, respectively). However, there was no difference between group obese non diabetic and control group as regard the allelic frequencies (p= 0.462). Activation of PPARg2 results in an increase in the sensitivity of both the liver to insulin-mediated suppression of hepatic glucose production and insulinmediated skeletal muscle glucose uptake. In addition, PPARg ligands are also shown to augment glucose disposal in peripheral tissues by increasing expression of the glucose transporter genes glucose transporter 1 (GLUT1) and GLUT4 (19). Our results showed that group IIIb had higher Pro allele frequency (p= 0.003) than in group IIIa (p= 0.048). This means that obesity could play an important role to make the effect of Pro 12 Ala polymorphism more pronounced in obese subjects. This may be due to the fact that PPARg2 gene is widely expressed in adipose tissue. So, negative mutation of this gene (as Pro 12 Ala polymorphism) is more manifest with more adipose tissue. Radha et al. (20) found that Pro allele is a genetic risk factor for type 2 diabetes mellitus in Caucasians but not in South Asians. Thus they suggested that population difference plays an important role in susceptibility to diabetes mellitus. Doney et al. (21) and Memisoglu et al. (22) also reported the association of pro allele frequency of PPARg2 and type 2 diabetes mellitus. Caramori et al. (23) showed similar results to our study and added that pro allele of this gene may predispose to diabetic nephropathy. In contrast to the current study, Mancini et al., 1999 (24) and Ringel et al. (25) failed to found a relationship between the pro 12 Ala polymorphism of PPARg2 gene and type 2 diabetes mellitus. Population difference may affect the role of Pro allele as a risk factor for type 2 diabetes mellitus by many theories such as the genetic background because of the genegene interaction or nutrition habits as some foods are considered PPARg ligands like oleic acid as reported by Soriquer et al. (26). Fasting blood glucose levels had no difference between Pro/Pro carriers and Pro/Ala plus Ala/Ala carriers in groups I, II and III (p= 0.17, 0.31 and 0.126; respectively). Doney et al. (21) and Tavares et al. (27) had reported similar findings as they found no difference between Pro/Pro carriers and Pro/Ala plus Ala/Ala carriers in control and diabetic groups as regard fasting blood glucose level. It is suggested that the PPARg2 gene affects the blood glucose level through enhancing insulin action on suppression of lipolysis, resulting in a decreased release of free fatty acids. Secondarily, reduced availability of free fatty acids would then permit muscle to utilize more glucose and liver to suppress glucose production more efficiently upon insulin stimulation. The non significant difference between Pro/Pro carriers and Pro/Ala plus Ala/Ala carriers in diabetic group could be explained by the fact that once diabetes has developed, the protective effect of the Ala allele may be lost because of the increased vascular complications and more pronounced β-cell dysfunction (28). Dubois et al. (29) suggested that the cause of β-cell dysfunction in Ala allele may be due to chronic exposure to elevated free fatty acids and/or hyperglycemia, which are characteristic for overt type 2 diabetes, leading to alteration or even reversion of the effect of the genetic variant. As regard the mean fasting insulin level there was significant increase in group II (obese non diabetic), group IIIa (diabetic non obese) and group IIIb (diabetic obese) when compared to group I (healthy control) (p= 0.001, 0.001 and 0.001; respectively) Also, the mean fasting serum insulin level was significantly increased in Pro/Pro carriers when compared to that of Pro/Ala plus Ala/Ala carriers (p= 0.39, 0.01 and 0.026; respectively in groups I, II and III). Our results agree with that of Ghoussaini et al, (3) and Tavares et al. (27) who had found significant increase in fasting serum insulin in Pro/Pro carriers when compared to that of Pro/Ala plus Ala/Ala carriers in obese group and diabetic group but not in control group. In type 2 diabetes mellitus, insulin secretion initially increases in response to insulin resistance in order to maintain normal glucose tolerance. The secretory defect initially is mild then it progress to a state of grossly inadequate insulin secretion. Insulin resistance is associated with obesity especially when centrally distributed, so obesity is accompanied with relative increase in serum insulin level to compensate insulin resistance. Also, chronic free fatty acids elevation impairs the insulin secretory response to glucose and, thus, plays an important role in the pathogenesis of diabetes (30). Ze-Kuan et al., 2006 (31) had reported that PPARg enhances glucose-induced insulin secretion as they found that overexpression of PPAR [gamma] in the absence of PPAR[gamma] ligands enhanced glucoseinduced insulin secretion by upregulation of both the glucose transporter 2 (GLUT2) and Cb1-associated protein (CAP) genes in β cells. Interestingly, Stevan et al. (32) found that lipid infusion designed to elevate plasma free fatty acids concentrations four folds resulted in a decrease in insulin secretion in carriers of the Ala allele, but an increase in control subjects with two Pro alleles. These findings might provide a partial explanation for the above observation that β cell function deteriorates more in Ala carriers once diabetes has developed. Dubois et al. (29) added for his explanation for the more deteriorated β cell function in Ala carriers once diabetes has developed, the superimposition of secondary mechanisms (such as chronic exposure to elevated free fatty acids and/or hyperglycemia, which are characteristic for overt type 2 diabetes) alters or even reverses the effect of the genetic variant. On the other hand, Stumvoll and Haring, (28) suggested that there is no strong evidence for a direct effect of the Pro12Ala polymorphism on insulin secretion. Also, this study showed that HOMA-IR was significantly increased in Pro/Pro carriers in group II and group III when compared to that of Pro/Ala plus Ala/ Ala carriers (p= 0.004 and 0.023; respectively) Our results coincide with that of Soriquer et al. (26) who had reported that Pro 12 Ala polymorphism of PPARg2 gene (pro allele) is associated with insulin resistance especially in population with a high intake of oleic acid and suggested an interaction between this gene polymorphism and dietary intake especially monounsaturated fatty acids which are considered as PPARg2 ligands. Scaglioni et al. (33) also were in agree with our results. They examined in their study the association between Pro 12 Ala polymorphism of PPARg2 and insulin resistance in obese children and found that Pro allele carriers had significantly increased HOMA-IR. Kao et al. (34) reported in their study that Ala allele of PPARg2 gene was associated with insulin sensitivity only among the nonobese individuals. They explained that the lack of association between the Ala allele and insulin sensitivity in obese individuals could be attributed to increased environmental influences, additional unknown genetic influences in this group, or the confounding influences of obesity itself. The explanation of the significant increase in insulin resistance in Pro/Pro carriers may be that Pro12Ala polymorphism of PPARg2 gene is a reduced function mutation (28). So it reduces the action of PPARg2 on insulin sensitivity through multiple mechanisms such as increasing expression of the glucose transporter genes glucose transporter 1 (GLUT1) and GLUT4. As regard the body mass index (BMI) the current study showed non significant difference between Pro/Pro carriers and Pro/Ala plus Ala/Ala carriers (p=0.58, 0.4 and 0.14 ; respectively in groups I, II and III). Our results coincide with that of Bouassida et al. (35) and Altshuler et al. (36) as they found no relation between pro 12 ala polymorphism of PPARg2 gene and BMI. However, Lie et al. (37) reported that pro allele of this gene was associated with higher BMI and body weight. On the other hand, Douglas et al. (38) demonstrated that pro allele is associated with lower BMI. The lack of association between pro 12 ala polymorphism of PPARg2 gene and obesity may be attributed to greater environmental influences, life style, gene nutrient interaction or additional unknown genetic influences in obese persons (27). The current study showed also that there was no difference between group II (obese non diabetic subjects) and group I (healthy controls) as regard Pro allele frequency (p= 0.462) these results agree with Ghoussaini et al. (3) Although PPARg2 gene is widely expressed in adipose tissue, its role in development of obesity is unapparent. The lack of association between pro 12 ala polymorphism of PPARg2 gene and obesity maybe attributed to greater environmental influences, life style, gene nutrient interaction or additional unknown genetic influences in obese persons (27). From this study, we can conclude that carriers of Pro 12 allele of PPARg2 gene could be more resistant to insulin than Ala 12 allele carriers and support the association between Pro allele of PPARg2 gene and type 2 diabetes mellitus. Thus, we can say that Pro 12 Ala polymorphism of PPARg2 gene may be a genetic risk factor for type 2 diabetes mellitus and its effect becomes more manifest when diabetes is accompanied with obesity. Further studies are recommended to investigate the effect of PPARg ligands in the treatment of patients with type 2 diabetes mellitus who carry the PPARg2 Pro 12 risk allele. Also, further studies are needed to clarify the role of this gene polymorphism in the development of obesity in order to help understanding the role of PPARg2 as an important regulator of insulin sensitivity and adipose tissue metabolism. So, will facilitate more effective strategy for management of insulin resistance. REFERENCES
Copyright 2009 - European Journal of General Medicine The following images related to this document are available:Photo images[gm09019t1.jpg] [gm09019t3.jpg] [gm09019t2.jpg] [gm09019f1.jpg] [gm09019t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}