 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Indian Journal of Human Genetics, Vol. 9, No. 2, July-Dec, 2003, pp. 55-64 Molecular phylogenetics and functional evolution of major rna recognition domains of recently cloned and characterized autoimmune rna-binding particle Erhan Süleymanoglu Institute of Biochemistry, University of Vienna, Medical Faculty,
Vienna Biocenter, Dr. Bohr-Gasse 9/3, A-1030, Vienna, Austria
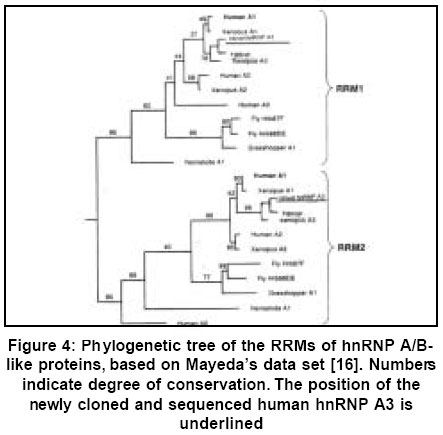
Code Number: hg03012 Heterogeneous nuclear ribonucleoproteins (hnRNPs) are spliceosomal macromolecular assemblages and thus actively participate in pre-mRNA metabolism. They are composed of evolutionarily conserved tandemly repeated motifs, where both RNA-binding and protein-protein recognition occur to achieve cellular activities. By yet unknown mechanisms these ribonucleoprotein particles are targeted by autoantibodies and hence play significant role in variety of human systemic autoimmune diseases. This feature makes them important prognostic markers in terms of molecular epidemiology and pathogenesis of autoimmunity. Since ribonucleoprotein (RNP) domain is one of the most conserved and widespread scaffold, evolutionary analyses of these RNA-binding domains can provide further clues on disease-specific epitope formation. The study presented herein represents a sequence comparison of RNA-recognition regions of recently cloned and characterized human hnRNP A3 with those of other relevant hnRNP A/B-type proteins. Their implications in human autoimmunity are particularly emphasized. Key Words: hnRNP proteins, RNA folding, RNA-protein interactions, Molecular evolution. Introduction The enclosure of eukaryotic genomes within the nuclear envelope evolutionarily generated the necessity to transport macromolecules selectively between nucleus and cytoplasm. Following their synthesis in cytoplasm, histones and nucleic acid polymerases, have to reach the nucleus, while specifically cytoplasmic proteins have to be kept out. Mature mRNA, tRNA and rRNA molecules and their associated proteins have to follow the opposite route, while their immature precursors have to be kept in.1-5 Eukaryotic mRNA is enzymatically metabolized and compacted with proteins within nuclei to generate functional messanger ribonucleoprotein (mRNP) particles.6,7 The control of protein biosynthesis involves regulation of intranuclear functions with participation of specific proteins, many of which seem to enter from the cytoplasm as a time specific events.3-7 The heterogeneous nuclear ribonucleoproteins (hnRNPs) comprise a group of important regulators engaged in these cellular processes.8-10. Extensive research efforts have been devoted to cloning and molecular characterization of numerous ribonucleoproteins in the context of their active role in mRNA biogenesis and metabolism,1,2,9-11 as well as in terms of their involvement as autoantigens in human diseases.11 RNA-protein interactions are considered as primary macromolecular forces governing gene expression. Hence, emphasis is put on the role of various RNA binding motifs and their subsequent role in governing cellular activities, both in health and in pathology.9 Recently, there has been considerable interest in the participation of these RNA-binding particles in triggering the immune responce in human autoimmune disorders.11 Substantial research both at cDNA and protein level have been performed on immunochemical features of human autoantigens (for continuously updated database see: http://www.zoo.uni-heidelberg.de/mol_evol/MB/ana_base.html). Of particular interest is the group of mammalian hnRNPs, which constitute a part of spliceosome, which itself is an autoimmune target.12 Novel nucleotide sequences are continuously reported, which afterwards are used to delineate homology among various members based on previously determined nucleotide and protein sequences. hnRNPs of diverse origin possess a common well-known structural motifs. However, the functional chemistry of these domains remains unknown. The mechanism of contribution of numerous hnRNPs with a similar structural RNP motifs13 to induction of different immune reaction is sequence specific.14 Phylogenetic approach is essential to find functionally important genomic sequences based on detection of their high degree of conservation across different species. Such approach shows the level of improvement of the prediction of gene-regulatory elements in the human genome. This necessiates the study of the degree of homology of RNA recognition motifs (RRM) among these proteins, requiring an evolutionary computation. Having considered the importance of submitting new sequences for further functional characterization, the newly cloned and expressed cDNA of previously unknown member of the hnRNP A/B family of proteins (Figure 1) is presented here. Its RNA-binding properties and tissue-specific gene expression profiles were recently determined.15 Based on the concept of correlation between sequences and RNA binding modes, a systemic search was performed for nucleic acid assosiation by evaluating sequence conservation by using multiple sequence alignments search tools. This study continues phylogenetic results obtained from previous larger data sets.16 Material and Methods 2.1. Preparation of RNA-binding domains To isolate the cDNA, human liver and brain cDNA libraries were screened by PCR using primers complementary to sequences in the 5'- and 3'-untranslated regions of the fetal brain (FBRNP) cDNA, as described.15 Screening of cDNA expression library resulted in isolation of a clone, which is a member of a 2xRNA-binding domain (RBD) - glycine family of hnRNP proteins. A cDNA was isolated from a human liver library encoding a 296 amino acid human hnRNP A3 polypeptide, highly homologous to the fetal brain cDNA, as well as to hnRNP A1 and A2. At the nucleotide level the highest degree of similarity is shared with a cDNA from Xenopus laevis. cDNAs encoding RNA-binding fragments of hnRNP A3 sequence were generated by PCR as deletion mutants, starting from the full-length human hnRNP A3. PCR mix was prepared by adding twice 660 μl water, 220 μl 10 mM dNTP mix (Pharmacia Biotech, Uppsala, Sweden), 220 ml 10x cloned Pfu polymerase buffer (Promega, U.S.A), and 122 μl 25 mM MgCl2. PCR primers used are shown on Table 1. 20 ml of each oligonucleotide (100 pmol/ml) was taken and added to 480 μl water. PCR reactions were run by taking 172 ml of the PCR mix (i. e., 86 ml per reaction), and mixing this amount with 12 ml of both primers. The amplification reaction was performed in a BiometraR, TRIO-Thermoblock™ PCR cycler. Denaturation was achieved by incubating for 30 sec. at 940C., while annealing temperature was 520C for 1 min. Extension reaction was performed at 720C for 1 min., 30 cycles. The amplified fragments were cloned into ligation independent cloning (LIC) vector (Novagen). 2.2. DNA Sequencing Clones containing the correct insert were sequences afterwards. All the DNA sequencing was performed by the Vienna Biocenter oligo team; http://emb1.bcc.univie.ac.at/gem. 2.3. Evolutionary analysis Nucleotide and deduced protein sequences of previously determined RNA-binding domains of ribonucleoproteins from various species and human hnRNP A3 were aligned employing CLUSTALW programme (http://www2.ebi.ac.uk/clustalw/),17 based on the experimental design of Mayeda, et al.16 Results The aim of the presented work herein was to isolate novel cDNA sequences with important functional implications in human pathology. Our efforts have been devoted to cloning and subsequent tissue-specific gene expression of numerous human ribonucleoproteins from the hnRNP A/B family of proteins (Figure 1).15 The objective was to search for molecular basis of autoimmunity by applying comparative analysis of the sequences of diverse autoantigens. In this context, evolutionary computation approach could give us major clues on how evolutionarily conserved mRNA transport machinery fails are linked to development of human autoimmune disorders. We were mainly interesting in cDNAs, which might encode the yet undescribed hnRNP B2. The need for this was based on two observations. Autoantibodies directed against hnRNP A2 crossreact with hnRNP B1 and hnRNP B2. Since hnRNP B1 is an alternatively spliced variant of hnRNP A2, this suggests that hnRNP B2 might be an alternatively spliced form of hnRNP A2/B1. However, all attempts to clone a cDNA encoding hnRNP B2 were not successfull so far. On the other hand, cDNAs closely related to hnRNP A1 and hnRNP A2 have been previously isolated from a human fetal brain library and from a Xenopus laevis library. Their close relationships with hnRNP A2 suggested that one of these cDNAs might actually encode hnRNP B2.15 To isolate the searched cDNA, human and liver and brain cDNA expression libraries were screened by PCR using primers complementary to 5'- and 3'-untranslated regions of the FBRNP cDNA. The isolated sequence seem to encode the full-length protein. Interestingly, however, it was not completely homologous to the FBRNP cDNA. Since the obtained new sequence shared close identity to the Xenopus laevis hnRNP A3 cDNA sequence (ENTREZ; accession number L02956), the protein was termed human hnRNP A3. Nucleotide sequence comparisons between FBRNP, Xenopus laevis hnRNP A3 and our newly determined human hnRNP A3 proteins revealed that extensive sequence conservation exist in RNA-binding regions. The differences observed here were mainly at the third position of the codon triplet. The majority of sequence variations were seen at the Gly-rich domain, composed of amino acids at positions 211-373. These were observed, as expected more at nucleotide level, as compared to the translated protein sequences (Figure 2). Only protein sequences are shown for brevity. Identification of various nucleic acid-binding domains of diverse hnRNPs was achieved by cloning and sequencing of cDNAs encoding these motivs. In general, all known human hnRNP proteins contain at least one RNA-binding modules and at least one another auxiliary domain fragment.The RNA-binding motifs contain the RNP consensus sequences (CS-RBD), the RNA recognition motif (RRM),10,13,19 the RNP-80 motif, the RGG box,20 and the KH domain.21 RNP domain is a the most common feature in these ribonucleoproteins. It is found in hnRNPs in varying amounts, ranging from 1 (in hnRNP C) to 4 (e. g. In Poly (A)-binding protein).10 Figure 1 shows the general modular structure of hnRNP A/B type of ribonucleoprotein particles. Their general structure is composed of two domains: the first 195 residues comprise the so-called UP1 domain, containing two canonical RNA-recognition motives-(RRM 1 and RRM 2), each of which is comprised from the conserved RNP-2 and RNP-1 submotives. The Gly-rich C-terminal domain comprises an RGG box and nuclear localization motif. This motif contributes to protein-protein interaction patterns, as well as to subcellular localization.20 Two conserved solvent exposed Phe residues, at the centre of the β-sheet in each RRM contact RNA. The least conserved 3rd loop in U1A is engaged in extensive RNA interactions, which conformation changes upon RNA binding.18-20 The RNP domain interacts with a flexible single-stranded RNA and the β-sheet provides a large surface for extensive interaction with nucleotides. Regions outside of the RRM may also play important roles in RNA binding.10 Identification of these motifs as RNA-binding domains has been used for prediction of this activity in various proteins of yet undescribed function possessing these domains. As seen (Figure 2), the newly isolated human hnRNP A3 cDNA encodes a protein of 296 amino acids. The calculated molecular weigh of 32 kDa was confirmed also electrophoreticaly. Amino acids numbered 1-98 comprise RBD1, amino acids 112-209 comprise RBD2, while the rest (amino acids 209-296) comprises the Gly-rich domain; amino acids 99-111 contain inter-RRM linker (IRL) segments. To our surprise, comparison with the previously reported FBRNP cDNA sequence, encoding a 269 amino acids protein, revealed only 85% identity. Therefore, it was assumed that the currently presented cDNA here, encodes a novel and yet undescribed protein. Despite the close homologies among the RNA-binding regions (95%), remarkable differences can be seen in the C-terminal domain (Figure 2), where deletions and insertions are apparent. Thus, the FBRNP is shorter by 27 amino acids and there are fewer conserved residues in the C-terminal part. Both cDNAs show also high homologies to the cDNA encoding hnRNP A3 from Xenopus laevis: except for a stretch of 14 amino acids at the RBD1 N-terminal part, the RNA-binding regions of the 3 proteins are almost identical, while the auxiliary domains are less conserved. The Xenopus laevis protein shows 75% homology with FBRNP and 83% with the newly cloned human hnRNP A3 protein, respectively (Figure 2). The reduced length of the human hnRNP A3, as compared to the Xenopus laevis homologue (296 versus 373 amino acids) is compatible with differences observed between Xenopus laevis and rat hnRNP A1, which are composed of 365 and 320 residues, respectively.15 The assumption that the presented clone encodes the human counterpart of frog hnRNP A3 is further reinforced, when conserved residues in the auxiliary domains of the A/B polypeptides are considered. Thus, a stricking homology is apparent at the C-terminus, which is well-conserved between frog and human hnRNP A3 with 15 of the last 18 amino acids being identical. Both, proteins end with a triplet RRF, which is also present in hnRNP A1, but neither in FBRNP nor in hnRNP A2/B1. Moreover, a glutamic acid located at the boundary of RBD2 and the auxiliary domains of Xenopus laevis and human hnRNP A3 (amino acid position 207) is substituted by valine in FBRNP, a further indication that the novel protein is indeed the human counterpart of Xenopus laevis hnRNP A3. Interestingly, this result is reported also for the murine hnRNP mBx protein, which is highly homologous to both human FBRNP and Xenopus laevis hnRNP A3.22 RRMs of hnRNP A/B type proteins share high degree of sequence homology. Most of the conformational differences between the two RRMs occur either at the C-terminal end of the αB helices, or at the tip of loop 3, which is tilted and twisted in RRM1, as opposed to RRM2 (Figure 3). Loop 3 is the least conserved region among different RRMs. The RNP domain interacts with a flexible single-stranded RNA and the β-sheet provides a large surface for extensive interaction with nucleic acids. The two aromatic side chains of RNP1 and one aromatic side chain of RNP2 provide a convenient template for base stacking of the RNA with neighbouring protein side chains, forming hydrogen bonds with the stacked bases. Although the major groove of the A-type RNA helix is too narow and deep to provide a site for sequence-specific association, an RNA loop with exposed nucleotides provides a large surface for protein binding. However, the RNA-binding is not unique feature of the β-sheets. Despite the evolutionary conservation of the RRMs, which are necessary for both general and sequence-specific nucleic aid-binding, regions outside of the RRMs may also play important roles in RNA binding.10 For instance, the flexibility of the linker sequence connecting both RRMs, resulting from two pairs of Arg and Asp involved in IRL salt bridges and the ordered residues creates a position for it, which is highly probable to be involved in direct RNA-binding. Figure 3. shows the RRM sequences of 12 hnRNP A/B type of proteins, extending the previous data set of Mayeda, et al.16 There is extensive sequence homology between these various proteins, which are conserved throughout evolution from insects to man. The sequences are 48-92% identical. Each of the RRMs forms an ungapped alignment with both RRMs of hnRNP A1. In all of the shown proteins, the RRMs are connected by a highly conserved IRL segments (Figure 1). The evolutionary links between the RRMs of human hnRNP A3 and other hnRNP A/B proteins are depicted in a phylogenetic tree (Figure 4). The sequences of RRM1 and RRM2 cluster in two separate groupings, representing insect and vertebrate proteins. The patterns of branching for RRM1 and RRM2 sequences are nearly identical, except for the placement of the placement of the branch representing the minor human variant hnRNP A0. The almost identical pattern of branching for both RRMs suggests that these fragments have evolved in parallel. The division of insect and vertebrate RRM sequences takes place on two separate branches. Obviously, each of the insect and vertebrate proteins are equally distant from each other. Apparently, this fact indicates independent gene duplications of ancestral hnRNP A/B-like protein, in agreement with Mayeda et al.16 Discussion The independent evolution of individual tandemly repeated protein domains and their functional relevance has always being an intriguing issue. The RNP domain is commonly encountered scaffold among the nearly 350 different folds, that is, the favourable secondary arrangements of arround 10 000 protein structures covered by the currently used databases.17,23-25 Comparisons of various protein structures, in combination with their nucleotide analyses serves as a clue for their evolution. The debate is whether these have evolved for achievement of specific functions or for thermodynamic stability and/or for kinetic folding reasons. The same structural topology can determine numerous activities, preserved through evolution. In biological systems, enzymes evolve by aquiring new thermodynamic or kinetic properties by the already existing protein folds. Generally, only the overall folding pattern is conserved in protein groups, and as the sequence diverge, the structures deform. Among the widespread catalytic folds, such as the TIM barrel, and the globins, the RRM domain attracts reasearch efforts into delineating the advancements of evolution starting from all α-helical proteins via all β-sheet proteins and reaching a group of α/β-proteins. In this route, the α/β-barrel is the most frequently seen enzymatic fold and appears to be a selected topology for the directed evolution of new biocatalysts. It is worth studying how the present a/b-barrels are linked evolutionarily to each other and through which way they have evolved from more simple ancestors. The topic becomes more attractive when these folds comprise a domain involved in a disease state, such as autoimmunity. The typical example here are the various hnRNPs, acting as autoantigens.9,11 The rapidly growing information on structure and function of numerous hnRNPs will be invaluable for understanding the pathological mechanisms by which these particles participate as targets of autoantibodies. Of particular interest is the determination of the role of RNA-protein folding patterns in development of disease-specific epitope formation. In this context, the evolution of selected nucleic acid-binding sequences and how this contributes to disease establishment remains unknown. Therefore, this study was devoted to presentation of evolutionary connections between the various hnRNPs and their RNA-binding domains. Phylogenetic tree is given to depict these motivs in the major group of these particles, including also the recently cloned and sequenced novel human hnRNP A3,15 followed by remarks on the lastly selected sequences, relevant in diseases. Figure 1 shows the general 2xRBD-Gly type of structure of the A/B type of human hnRNPs. The two canonical RRM-1 and RRM-2, containing the conserved RNP-2 and RNP-1 submoieties, and the RGG C-teminal part constitute the main domains. The translated sequences of the recently characterized human hnRNP A3 particle (Figure 2) is presented herein, solely to demonstrate the high degree of sequence homology among the different RRMs. Its protein sequence is compared to that of other two members of this protein family -Xenopus laevis hnRNP A3 and FBRNP for brevity. More detailed motif comparisons are given afterwards (Figures 3 & 4). The selected three members at Figure 2 are functionally related quite distantly from each other, each having a buried core tryptophan and there are disulphide bonds on the highly conserved IRL-segments.16 Usually, conservation of activity is not expected, because of the evolutionary pressure. The sequence comparisons of the three closely related hnRMP A3 members showed that the general need for a large buried hydrophobic moieties is well satisfied. This is evolutionarily highly conserved due to thermodynamic stability requirements. This represents a possible case, where structural constarints make evolution towards a different conformation unlikely. The high level of conservation of these residues is related to the interlocking interactions and restraints of the loop connecting the β-sheets, by providing the huge hydrophobic template necessary for core packing. In addition, having large hydrophobic inner part and suitable hydroxyls capable of hydrogen-bond formation, these residues (e.g. tyrosines) could be conserved for structural stability needs. Performing amino acid mutations, or substitutions at the sites thus removing hydrophobic packaging interactions would provide further evidence for the core packing of hnRNP globule. Moreover, the role of other conserved residues is worth to approach. These are buried in the compacted core and are expected to participate in overall forlding of the scaffold. However, to verify whether this is the case, more mutational analyses, combined with thermodynamic, kinetic, structural and evolutionary computation data are needed. Amino acid sequence comparisons of RRM structures (Figure 3) depicted the hydrophobic core residues, conserved among different RRMs. The inserted sequence of the human hnRNP A3 shared extensive sequence homology (up to 90%) with RRMs of all these numerous proteins, which are conserved throughout evolution. Phylogenetic tree (Figure 4) depicted the evolutionary history of the RRMs of human hnRNP A3 and other hnRNP A/B proteins. The objective was to deduce whether the observed patterns of conservation resulted due to structural features for this type of fold architecture, or alternatively, are the consequence of divergent evolution. Parallel symetric groupings of RRM1 and RRM2 in two different clusters, to which belong insect and vertebrate members is well seen. With the slight deviation of hnRNP A0, the rest of the RRM branch sequences are almost identical. This branching pattern is an indication of parallel proceeding evolution, in accordance with relevant studies.16,27 Each of the members of these two subgroups are equally distant from each other. These results suggests that these branching patterns originated from an ancestral hnRNP A/B-like protein probably though independent gene duplications. Phylogenetic analyses of this sort provide evidence for evolutionary history and origin of the modular structure of hnRNPs and their contemprory functional implications. This approach helps to delineate whether RNA-binding (RBD) and protein-protein (auxiliary) domains have evolved in parallel or follow another evolutionary history. In this context evolutonary computation on every newly characterized hnRNP particle become valuable and a prerequisite for understandanding of its nowadays functions. The sequence of human hnRNP A3 were inserted in the data set at Figure 4 and becomes a further support of evolutionary trend of these ribonucleoprotein family of proteins, appearing also in previous studies.16,27 These earlier studies proved that the origin of hnRNPs is a consequence of independent gene duplication. In their landmark work, Fukami-Kobayashi et al. 27 showed that the ancestral gene of the hnRNPs had had two RNA-binding domains even before the divergence of invertebrates and vertebrates and that it diverged to contemporary hnRNP genes while preserving the tandemly repeated structures. The origin of natural selection of particular nucleotide sequences and their conservation from primitive RNA based catalysis to eukaryotic chromosome evolution remains to be elucidated.28-30 In the light of "primordial RNA catalysis doctrine",28 there is now sufficient data to prove the substitution of ancestral RNAs by more efficient RNA-protein complexes with subsequent replacement of the latter by proteins.29 By using molecular modeling approach, it has been recently shown30 that in this transition position, the two RNA-binding sites of the simulated proto-protein interact with target sites to form a stable RNP complex. Since the selected proteins were unable to resolve misfolded RNA, in vivo selected protein associated with the two RNA-binding sites, indicating that the protein facilitates the correct folding of the ribozyme. This provides evidence that these modelled self-splicing RNA-protein complexes can be considered as a primitive form of splicing factors or RNA chaperones, which contribute to correct RNA folding.30 In terms of molecular evolution of primitive catalysts,28 these self-splicing intron assemblies possibly became splicing RNPs, suggesting that an active RNP particles could have originated from a primordial ribozymes.30 The evolutionary driving force for this is the small structural cost, through which protein binding compensates for some folding deficiencies in the RNA.31 This serves as another proof that the evolution from RNA to RNP-determined catalysis represents an evolutionary design against misfolding rather than for maintanance of a protein-binding site. Our preliminary biophysical results26 suggests that this is the case with human hnRNP A3, as well. The current study presents the sequence conservation of mainly hydrophobic residues of human hnRNP A3, which form the hydrophobic packing cores and are essentially evolutionarily conserved in other RNA-binding domains in the hnRNPs studied (Figures 2-4). This indicates that the RNA-binding domains have evolved from a common hnRNP A/B-type proto-protein, supporting further previous models.16,27 Interestingly, while the majority of models of RBD protein family built up to now are based on divergent evolutionary principles, another view claims that this is open to doubt in cases of similar, but more distantly related functions and structures.32 Their model suggests that even though the ancient RBDs could have developed separately by divergent evolution, these have evolved conserved RNP motivs with a similar structure and function on similar surfaces. The authors emphasize that RNP motifs are conserved both in scaffold architecture and in function, which provides an intriguing case of convergent evolution. Further advancements in sequence comparison techniques will help to understand whether these functional sites have arisen multiple times during evolution.17,23,24,33,34 Regarding the evolutionary conservation of hnRNP domains two possibilities exists for explaining of their nowadays existance-they are either hyperadaptable, or they may have developed features required to perform vital cellular functions.33 Despite the case with cold shock domain,32 hnRNP A/B proteins appear to possess eukaryotic origins, because prokaryotic homologs could not be detected until now. All these RBDs have similar dimerization roles leading to similar three-dimensional architectures, suggesting that they may have arisen from a common ancestor, which have diverged in sequence afterwards.33 These domains are found also in chloroplast protein sequences, implying the link with endosymbiont hypothesis and that multicellularity in plants and and animals did not evolved independently.27,32,33 Domain evolution of hnRNPs rises the question of whether they are consequences of continuous or discontinuous evolution. Go M, et al27 showed that RBD have fused with the SR-rich domain preceeding the divergence of splicing factors, and that these two domains have arisen together thus conserving the fused domain organization. The existance of Gly-rich domain (Figure 1), also supports the proposal of domain organization of the unique origin of hnRNPs, followed by duplication and divergence of RBDs and structural transitions of the auxiliary domain.20 This indicates that the auxiliary domain shared by functionally related ribonucleoproteins diverged in parallel together with the RBDs.27 It is thus implied, that these ancestral repeats must have oligomerized afterwards to adopt a similar structures seen in contemprory homologs.33 In the light of the fact that hnRNP A/B particles are considered as disease genes, an intriguing issue becomes the use of protein domains data of various hnRNPs to delineate the link to and etiology of diseases. As stated earlier, major of these gene activities related to autoantigenic hnRNPs are covered by relevant databases. These are useful for prediction of functions of newly sequenced pathogenic genes. Within this respect, identification of human paralogous disease genes generates further pathological gene candidates to be sequenced, because paralog genes are frequently encountered as mutated versions in similar diseases.33 The sequence comparison, however, may be insufficient for deduction of its functional role for patients diagnozed with the suspected disease. Therefore, we followed the suggestion of Ponting CS And Russell RR.33 and combined our sequence determination of human hnRNP A3 with its gene expression profiles, its molecular interaction features,26 and its tissue-specific gene expression patterns.15 Interestingly, our molecular characterization of human hnRNP A3 showed that while the recombinant hnRNP A3 with its 296 amino acids migrates as expected as 32 kDa protein on SDS-PAGE analysis, it is recognized by the patients' sera as a 50 kDa protein. This is attributed to alternative splicing or due to tissue-specific expression of a highly related yet unknown crossreactive protein. Surprisingly, neither the 50 kDa, nor the 32 kDa protein was detected in He Lanuclear extracts further supporting the assumption that hnRNP A3 is not a ubiquituously expressed as hnRNP A1 or hnRNP A2, respectively. Northern Blotting analysis showed the variable expression patterns of hnRNP A3 mRNA in human tissues. Thus, it is highly expressed in spleen, ovary, small intestine, lung, liver, skeletal muscle, kidney and pankreas, whereas expression in thymus, testis, colon and peripheral blood was hardly detectable. Expression in prostata, heart, brain and placenta was low, but clearly detectable.15 In addition, circular dichroism and fluorescence spectroscopic measurements demonstrated that the human hnRNP A3 protein is a stable particle - the free energy of unfolding of the full-length hnRNP A3 is 580C, as shown by both urea and temperature denaturation. 26 This particle increases its apparent stability upon interaction with RNA fragments r(UUAGGG)4 to higher temperature values. The high binding affinity to this repeats indicates its preference for association with purine-rich consensus sequences and targeting features towards deleterous G-tetrad structures. The latter fact suggests its active participation in alternative splicing-a common feature of well known telomeric repeat interactions of hnRNP A1. This RNA fragment-hnRNP A3 particle recognition can act by facilitating the splicing of alternative intron of the pre-mRNA. Since this represents a case of regulation of splice-site selection with further functional implication in human disease,11,12,35,36 epitope mapping studies of human hnRNP A3 were carried out. HnRNP A3 possesses two majour autoepitopes- first comprised of both RBD1 and RBD2 and the second one, composed of RBD2 and certain parts of Gly-rich domain.15,26 This epitope recognition pattern differs from the epitope determined for the highly related hnRNP A1 and hnRNP A2, respectively. Thus, in hnRNP 2 the RBD2 was found to contain the major epitope, which was recognized by patients suffering from rheumatoid arthritis or systemic lupus erythemathosus. This particular region was also found to be essential for interaction with RNA and the patients' autoantibodies strongly inhibited RNA binding. As opose to these, autoantibodies derived from patients diagnozed with mixed connective tissue disease recognized an epitope comprising RBD1 and RBD2. Interestingly, the major epitope of hnRNP A1 also comprise both RBDs, which is, however, targeted also by patients with rheumatoid arthritis and systemic lupus erythemathosus. Taken together, these data confirm that rheumatoid arthritis, systemic lupus erythemathosus and mixed connective tissue disease are immunochemically linked by systemic autoimmunity to the functionally important RNA-binding regions of hnRNP A/B proteins. The observed trend brings the question of why are so closely related and evolutionarily conserved RBD domains differentially recognized by the autoantibodies. We proposed recently26 that the overall folding patterns of these domains and the protein antigenicity arising from this is the determining factor. The present study combined with previously determined sequences of U1A, hnRNP C, hnRNP A1 and Drosophila sex lethal (sxl) protein structures reveals that they have the same fold, but different placing of the second and fourth ß strands. Individual RRMs have preferences towards various RNA sequences, due to differences in surface amino acids found outside the conserved RNP submotifs. In our opinion, this can explain the differential recognition of RRMs by patients' autoantibodies. In certain cases the two RRMs somehow act in concerted fashion to give rise to the overall RNA- and antibody-binding characteristics, whereas in other cases the presence of only one RRM is sufficient for autoantibody and RNA recognition. Thus, RBD1 itself bound strongly to RNA fragments, however, joining of RBD2 to RBD1 increased the overall affinity indicating that RBD2 also strongly affects RNA-binding. This evidence indicates the possible existance of RNA conformation-specific autoantibodies. Aknowledgements I thank Dr. Richard Hrabal (Laboratory of NMR Spectroscopy - Institute of Chemical Technology, Prague, Czech Republic: http://www.vscht.cz/nmr for the hospitality and generous financial support during my stay in his laboratory in Prague as a guest researcher and for the detailed supervision concerning our joint work on NMR studies of RNA-protein recognition and its significance in the control of stability of hnRNP particles. References
Copyright 2003 - the Indian Society of Human Genetics The following images related to this document are available:Photo images[hg03012f2.jpg] [hg03012f4.jpg] [hg03012f1.jpg] [hg03012t1.jpg] [hg03012f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}