 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
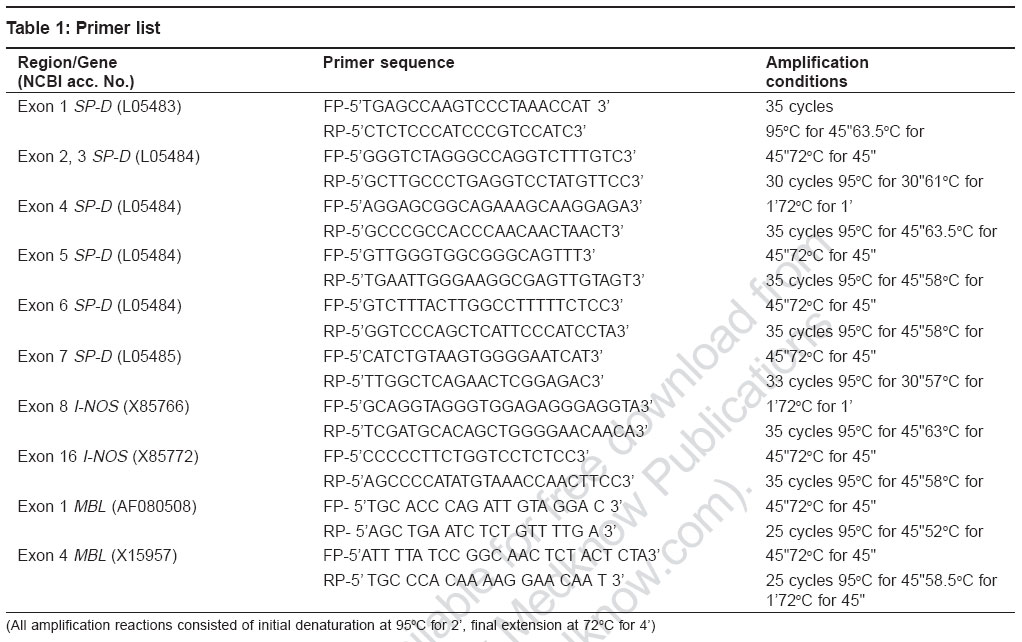
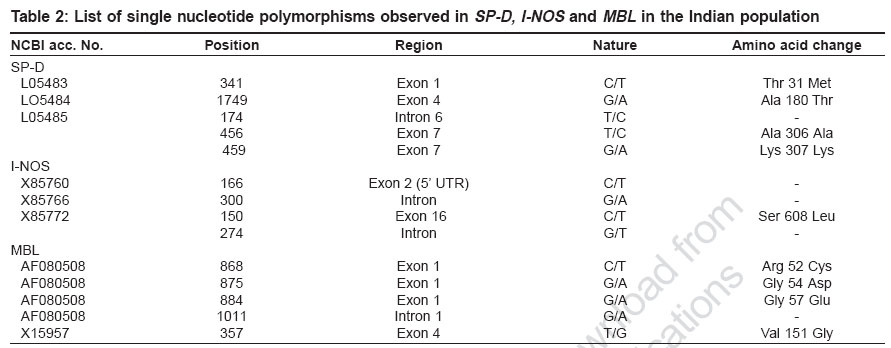
Indian Journal of Human Genetics, Vol. 12, No. 3, September-December, 2006, pp. 105-110 Original Communications Association of SP-D, MBL and I-NOS genetic variants with pulmonary tuberculosis Vaid Mudit, Kaur Savneet, Taruna Madan, Singh Hari, Gupta VijayK, Murthy KJR, Sarma PuranamU Molecular Biochemistry and Diagnostics, Institute of Genomics and Integrative Biology, Mall Road, Delhi Code Number: hg06019 Abstract Background : Pulmonary tuberculosis is caused by Mycobacterium tuberculosis . It is a multifactorial disease with both host as well as pathogen factors contributing to susceptibility and protection from the disease. Various reports have highlighted important roles of lung surfactant protein D (SP-D), mannan-binding lectin (MBL) and I-NOS in innate immune defense against M. tuberculosisAims : The present study investigated the role of polymorphisms in three candidate genes encoding Lung surfactant protein D, Mannan binding lectin and Inducible Nitric oxide synthase, in susceptibility and protection to pulmonary tuberculosis. Settings and Design : A case-control association study of SNP's in lung surfactant protein D (SP-D), mannan-binding lectin (MBL) and I-NOS with pulmonary tuberculosis in Indian population was carried out. This involved sequencing of all the coding exons of lung surfactant protein D (SP-D) , while, exon 1 (collagen region) and exon 4 (carbohydrate recognition domain) of mannan-binding lectin (MBL) and exons 2, 8 and 16 of I-NOS and their flanking intronic regions for single nucleotide polymorphisms in DNA samples isolated from 30 pulmonary tuberculosis patients and 30 controls of Indian population. Statistical analysis: Various allele frequencies were calculated using online two by two table (home.clara.net/sisa/). Odds ratio and P values were calculated at 95% confidence interval (CI). Results : A total of fourteen single nucleotide polymorphisms (5 in SP-D , 5 in MBL and 4 in I-NOS ) were observed of which four (G459A SP-D , G274T I-NOS , G1011A and T357G MBL ) have not been reported earlier. Four single nucleotide polymorphisms viz. G459A of exon 7 of SP-D ( P =0.00, odds ratio (OR) = 4.96, 2.18 Conclusion: The present study has led to identification of 4 SNP's in SP-D , MBL and I-NOS associated with pulmonary tuberculosis in Indian population. Keywords: Inducible nitric oxide synthase, lung surfactant protein D, mannan binding lectin, polymorphism, pulmonary tuberculosis Introduction M. tuberculosis is a ubiquitous pathogen: estimates are that roughly one third of the world′s population is infected with the bacillus and it is responsible for eight to 12 million cases of active tuberculosis each year and three million deaths.[1] There is compelling clinical evidence that, cross-talk between bacterial pathogenicity and host immune response against M. tuberculosis determines the clinical manifestations and ultimate outcome of the infection.[2],[3] Collectins such as, lung surfactant proteins and mannan-binding lectin (MBL) have evolved as key players of innate immunity against various pathogens,[4],[5],[6],[7],[8] including mycobacterium . Several studies suggest an opsonic role for lung surfactant protein A and MBL against mycobacterium .[9],[10],[11] However there are conflicting reports regarding role of Lung surfactant protein D (SP-D) in mycobacterial opsonization.[12],[13] Suggesting a modulatory role for SP-D in mycobacterial phagocytosis, with exact role of SP-D still evading. Besides binding and opsonization, a role of macrophage activation has also been assigned to the collectins which leads to enhanced phagocytosis and killing of mycobacteria through a concerted activity of IFN-g and TNF-a which operates through participation of reactive oxygen intermediates (ROI) and reactive nitrogen intermediates (RNI).[14] RNI and especially NO have been implicated as important defense molecules against mycobacteria .[15],[16],[17] Out of two NO producing enzymes namely inducible and constitutive (endothelial and neuronal) nitric oxide synthases, NO produced by inducible nitric oxide synthase (I-NOS) is relevant for innate immunity against pathogens. Since, the incidence of TB is higher in immunocompromised individuals,[18],[19] studies on innate immune factors are of particular significance. We believe that the long arm of Chromosome 10 which houses SP-A, SP-D and MBL in close proximity is an ideal candidate region for screening genetic factors associated with tuberculosis. In our earlier studies, we observed association of polymorphisms of SP-A2 with pulmonary tuberculosis in Indian population.[20] The association of SP-D polymorphisms with TB has been reported in Mexican population.[21] In case of MBL, there are conflicting reports regarding association of ′B′ allele of MBL (codon 54 polymorphism in exon 1, coding for low serum MBL levels), with pulmonary tuberculosis in different populations.[22],[23],[24],[25],[26] The 17q11-q21 region, housing I-NOS gene, was mapped as susceptibility locus for tuberculosis in Brazilians.[27] So in the present report, we have carried out case-control association studies of SNP′s in SP-D , MBL and I-NOS with pulmonary tuberculosis in Indian population. This involved sequencing of all the coding exons of SP-D, while, exon 1 (collagen region) and exon 4 (carbohydrate recognition domain) of MBL and exons 2, 8 and 16 of I-NOS and their flanking intronic regions for single nucleotide polymorphisms since these regions have been shown to be functionally important in earlier studies. Materials and Methods Study design Following approval of the Institute′s Human Ethics Committee, SNP screening of coding exons and flanking intronic regions of SP-D, MBL and I-NOS was carried out in Indian subjects. The study comprised of group of 30 patients of pulmonary tuberculosis enrolled at the Mahavir Medical Research Center, Mahavir hospital, Hyderabad, India. Diagnosis of TB was based on demonstration of acid-fast bacilli (AFB) in sputum and was eventually confirmed by positive culture of Mycobacterium tuberculosis. A second group of 30 healthy, tuberculin non-reactors, age and sex matched to patients was taken as the control. Genomic DNA isolation Human genomic DNA was isolated from blood using modified salting out procedure.[28] Briefly nuclei were isolated from 10 ml blood (collected in ACD) by addition of ice-cold cell lysis buffer to the blood followed by centrifugation at 3100 rpm for 15 minutes. To the pelleted nuclei then added 12 ml of nucleus lysis buffer, 0.8 ml of 10% SDS and 50uL of proteinase K (20 mg/ml). Proteins were then precipitated using 4 ml of 6M NaCl and finally pelleted by spinning at 2500 rpm for 15 minutes. To the supernatant added exactly two volumes of room temperature 100% ethanol and inverted the tube several times until the DNA precipitate is visible, which was then removed with a plastic spatula or pipette tip and transferred to an eppendorf tube containing 100-200 ml TE. Polymerase chain reaction (PCR) All PCR amplification reactions used 50 ng of the template DNA, 1.5 mM MgCl 2 , 0.2 mM dNTP ( Amersham, UK ), 1.5 units of Taq polymerase ( Bangalore genei, India ) and 20.0 pico mole each of forward and reverse primers, in 50 mL of reaction. Primers for Exon 2 of I-NOS were taken from previous report[29] while [Table - 1] summarizes the sequence and amplification conditions for other primers used in the present study. Purification of the PCR products PCR products were purified by PEG-8000/Sodium acetate (3M, ph 4.8) precipitation except in the case of exons 1 and 4 of SP-D where purification was carried out using DNA elution Kit (Cat no. 20-200-300, Biological Industries) as per manufacturer′s instructions. Purified samples were air dried and resuspended in 10 mL of autoclaved MilliQ water and quantitated on 1% agarose gel. Cycle sequencing Purified PCR products thus obtained were subjected to cycle sequencing reaction based on dideoxy termination reaction. Briefly 35-50 ng of the purified PCR product was amplified using 1pico mole of the forward primer and 4.0 μL of ready reaction mix in a 10 μL reaction. Autoclaved MilliQ water was used to make up the volume wherever required. Amplified product was precipitated using 150 mM EDTA and 3M NaOAc followed by two 70% ethanol washes and air-dried. Samples were loaded on capillary-based ABI 3730 sequencer as per the standard procedures. Cycle sequencing program consisted of 25 cycles of denaturation at 96°C for 10 seconds, annealing at 50°C for five seconds, extension at 60°C for four minutes and final incubation at 4°C. Sequence data obtained was analyzed for the polymorphisms using basic local alignment search tool (http://www.ncbi.nlm.nih.gov/BLAST/) and Seqman software of DNA star. Statistical analysis Statistical analysis and frequencies of various alleles were calculated using online two by two table (home.clara.net/sisa/). Odds ratio and P values were calculated at 95% confidence interval (CI). Results In the present study we screened polymorphisms in all the coding exons of SP-D , exons 1 and 4 (encoding collagen and CRD region respectively) of MBL and exons 2, 8 and 16 of I-NOS and investigated the role of these polymorphisms in genetic predisposition to pulmonary tuberculosis in Indian population. We found that at a number of places (21 in SP-D , 25 in I-NOS and 1 in MBL ) the observed sequence did not match the sequence reported in the database. We have assigned them as changes [data not shown] and not polymorphisms as these changes were observed in all the samples analyzed in the study irrespective of their health status. Interestingly, all but three (2 in SP-D and 1 in MBL) changes were intronic. A total of fourteen single nucleotide polymorphisms ([Table - 2]; 5 in SP-D , 5 in MBL and 4 in I-NOS ) were observed in the present study. To the best of our knowledge four of these SNP′s (G459A SP-D , G274T I-NOS , G1011A and T357G MBL ) have not been reported earlier. A comparison of allele frequencies between controls and TB patients [Table - 3] revealed that ′A′ allele at position G459A of exon 7 of SP-D ( P = 0.00 or= 4.96; 2.18 < OR< 11.31) was significantly associated with TB in Indian population. Interestingly, none of the subjects screened was homozygous for ′A′ allele. The mutant 'A' allele of G875A in codon 54 of MBL exon 1 (P=0.00 or =3.85 1.66OR<8.88) showed significant association with TB in the Indian population. The frequency of mutant allele of MBL codon 57 ('A' allele G884A), was similar in patients and controls while mutant allele for MBL codon 52 ('T' allele of C868T), was observed only in one of the controls. The movel SNP T357G in codon 151 of MBL exon 4 involved a change from Valine to Glycine. However, the frequency of mutant 'G' allele of T357 G was similar in patients and controls and hence showed that two patients were homozygous, whereas we didn't detect the presence of any functional mutant homozygote of other SNPs in exon 1 and exon 4 of MBL. In I-NOS, G allele at position G300A of intron 8 (P=0.00 or=4.04, 2.20 < OR < 7.42) is associated with tuberculosis with frequency of homozygous 'GG' patients almost 2.5 times that of homozygous controls. T allele at position G274T of intron 16 of I-NOS (P=0.00 or=4.46, 2.40 < OR < 8.28) showed significant association with pumonary TB in Indian population [Table - 3]. Genotype patterns for patients and controls w.r.t G274T revealed that none of the controls were homozygous for the mutant 'T' allele, whereas five out of 20 (25%) patients showed homozygosity for this allele. Discussion Association of TB with alleles of various genes like vitamin D receptor, interferon-gamma, mannose-binding lectin, IL-1 and NRAMP etc, suggest it to be a multifactorial disorder with various host genetic factors regulating susceptibility and/or protection against the disease.[30],[31],[32],[33],[34],[35],[36] ′A′ allele at position G459A of exon 7 of SP-D was found to be significantly associated with TB in Indian population. This is a novel but synonymous polymorphism, with both the alleles coding for lysine. Because of the polygenic nature of TB, there is a possibility of A allele of G459A being in linkage disequlibrium with some other important genetic factors. Earlier Floros et al have identified DA11_C in exon 1 of SP-D as susceptibility locus for tuberculosis in Mexican population.[21] This polymorphism was not observed in the present study. Further our data on genetic polymorphisms in SP-D in Indian population suggests that SP-D, unlike SP-A, is highly conserved. In case of MBL , a significant association of ′B′ allele with tuberculosis is similar to the earlier observations made by Selvaraj et al in Indian population.[22] A similar observation in African-Americans has also been reported.[23] MBL forms the humoral mediator of mycobacterial phagocytosis and a reduction in the levels of functional MBL in the serum caused by the presence of variant alleles of MBL may affect the initial defense against M. tuberculosis It has been suggested that the presence of ′B′ allele, which leads to low levels of MBL in the serum, might affect the antigen presentation by macrophages and antigen recognition by lymphocytes, which may lead to decreased lymphocyte response M. tuberculosis antigen.[24] However our observations are not in coherence with those of Soborg et al in Caucasians[25] and Ozbas et al in Turks.[26] Sequencing of I-NOS revealed that susceptibility to infection is associated with two intronic SNP′s in I-NOS namely G300A (intron 8) and G274T (intron 16). Since G300A is only 12 bases downstream of exon 8 - intron 8 boundary, a site for alternative splicing[29] it may be affecting the splicing. C166T polymorphism in exon 2 (coding 5′UTR) and C150T, a nonsynonymous exonic polymorphism (Ser to Leu) in exon 16 of I-NOS already reported in the Danish population,[29] were observed in the Indian population but did not show association with pulmonary tuberculosis in the Indian population. The present study has led to identification of 4 SNP′s in SP-D , MBL and I-NOS associated with pulmonary tuberculosis in the Indian population and has enriched the already known list of genetic markers for predictive diagnosis of tuberculosis. Acknowledgments This work was supported by the Council for Scientific and Industrial Research (T.M, P.U.S.) (CMM0016). We acknowledge the technical expertise provided by The Center for Genomic application (TCGA), a collaborative effort of the Institute of Genomics and Integrative Biology, (a CSIR organization), supported by DST and Institute of Molecular Medicine (a Chatterjee Group company) for the sequencing of PCR products. Mudit Vaid and Savneet Kaur are recipients of Senior Research Fellowships, Council of Scientific and Industrial Research. References
Copyright 2006 - Indian Journal of Human Genetics The following images related to this document are available:Photo images[hg06019t1.jpg] [hg06019t2.jpg] [hg06019t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}