 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
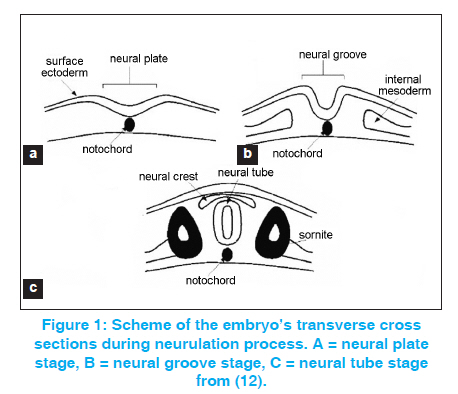
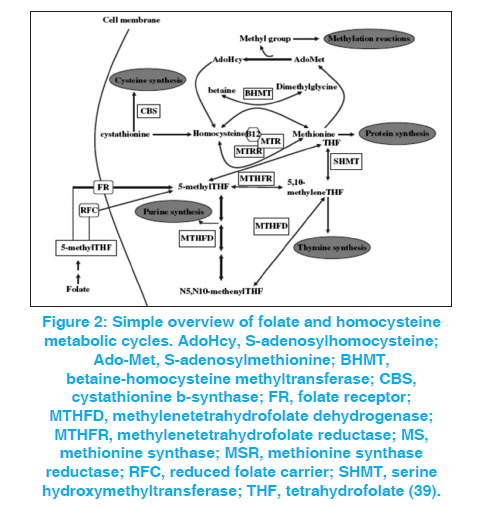
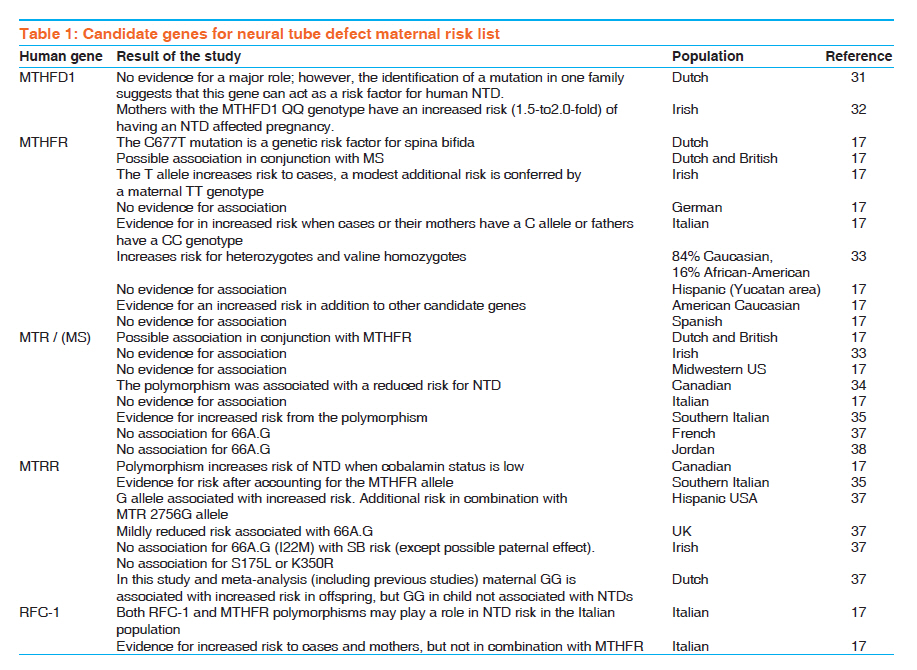
Indian Journal of Human Genetics, Vol. 17, No. 3, September-December, 2011, pp. 126-131 Review Article Neural tube defects between folate metabolism and genetics Helmi Y Alfarra, Sabreen R Alfarra, Mai F Sadiq Department of Biological Sciences, Yarmouk University, Irbid, Jordan, Code Number: hg11031 DOI: 10.4103/0971-6866.92082 Abstract Neural tube defects (NTDs) are the second most common severely disabling human congenital defects. Worldwide, NTDs incidence is approximately one per 1000 live births and varies between 0.78 and 12 per 1000 births in general populations. NTDs causation involves multiple genes, nutritional and environmental factors. Research in the next stage should include bigger populations and bigger studies that would be suitable to detect significant links and conclusions with relatively minor risk factors; analysis of multiple candidate genes to detect potential gene-gene interactions; detailed analysis of patient samples stratified by phenotype. Keywords: Folic acid, MTRR, MTR, MTHFR, NTDs Introduction The neural tube is the embryonic precursor of the brain and the spinal cord. Building a neural tube is an extremely complex phenomenon where cells need to change in shape, migrate and differentiate to form a hollow tube from a flat sheet of thickened epithelial cells (the neural plate). The normal closure of the neural plate occurs very early in human pregnancy from day 21 until day 28 following conception or one week after the expected menstruation. [1] Abnormal closure of the neural plate results in neural tube defects (NTDs). NTDs are very complex birth defects that involve the interactions of multiple systems such as the central nervous system, the urological system and the musculoskeletal system. NTDs seem to be related to defects in the folate-methionine metabolic pathways since preconception of vitamin B12 supplementation with folic acid proved to substantially reduce the risk for NTD affected pregnancies and many different investigations showed alteration in folate status in the mothers of NTDs affected children. In these mothers, folate absorption, transport, and metabolism are the focus of intense research to clarify the mechanisms underlying these observations, yet the bases of this defect remain unknown. Further studies involved examining the different polymorphisms of genes related to folate metabolism. [2],[3] NTDs are the second most common severely disabling human congenital defects. Worldwide, NTDs incidence is approximately one per 1000 live births and varies between 0.78 and 12 per 1000 births in general populations. [4],[5],[6] In Jordan, three studies were performed during the years 1991 to 2003, showed that the average incidence of NTDs in the Jordanian population is about 3.8 per thousand. [7],[8],[9] The correlation between the birth of NTD affected children and the maternal polymorphisms of two genes is related to the folate metabolic pathway. Methylenetetrahydrofolate-dehydrogenase (MTHFD1) 1958 (G>A) and methionine synthase polymorphisms MTR 2756 (A>G) and MTR 2758(C>G) in the Jordanian population have been examined. Development of the neural tube (neurulation) Neurulation is a fundamental embryonic process that leads to the development of the neural tube, which is the precursor of the brain and spinal cord. Neurulation occurs through two distinct phases: primary neurulation (weeks 3-4) that leads to the formation of the brain and most of the spinal cord till the upper sacral level, followed by secondary neurulation (weeks 5-6) that creates the lowest portion of the spinal cord including most of the sacral and all the coccygeal regions. [10],[11] Primary neurulation This process involves formation of the neural plate, shaping of the neural plate, bending and fusion of the neural plate at the midline. [12] Shaping of the neural plate involves conversion of the neural plate into an elongated structure that is broad at the cranial and narrow at the spinal regions. In this complex process, cells elongate mediolaterally and produce polarized cellular protrusions that enable them to move directionally and to intercalate with other neighboring cells. This change in shape and movement results in convergence toward the midline and extension of the tissue along the anteroposterior axis. Bending of the neural plate implicates formation of the neural folds at the lateral extremes of the neural plate and the subsequent elevation and convergence of these folds toward the dorsal midline [Figure - 1]. [12] Secondary neurulation Secondary neurulation begins after completion of the primary neurulation. During this process, the neural tube is produced by the tail bud, a mass of stem cells representing the remnant of the primitive streak that are located at the caudal end of the embryo. These stem cells undergo proliferation and condensation followed by cavitation and fusion with the central canal of the neural tube that is formed by primary neurulation. [13],[14] Failure of secondary neurulation leads to the rare closed forms of NTDs where the developing neural tube fails to separate from other tissues of the tail bud. [14] Folate metabolism Folate is a critical nutrient in maintaining normal cell growth and division and therefore it is important for human development. [15],[16] After uptake of folate in the enterocytes, intracellular polyglutamate synthesis occurs, followed by reconversion into monoglutamates that are reduced to biologically active tetrahydrofolate. [16] After methylation, the folates enter the portal circulation as 5-methyltetrahydrofolate. The majority of this folate is taken up by the liver, which plays a central role in folate homeostasis. [16] The cellular uptake is mediated via specific folate receptors. Plasma 5-methyltetrahydrofolate (30-40%) is bound to albumin, α2 macroglobulin, transferrin, and folate-binding protein. [15] In the cell, 5- methyltetrahydrofolate serves as a methyl donor and as a source of tetrahydrofolate. Tetrahydrofolate acts as an acceptor of one-carbon units, producing a variety of other folates, which, in turn, are specific coenzymes in intracellular reactions. [16] One of these is 5, 10- methylenetetrahydrofolate that is reduced by the enzyme 5, 10- methylenetetrahydrofolate reductase to 5-methyltetrahydrofolate. 5-Methyltetrahydrofolate carries the methyl group required for conversion of homocysteine to methionine (remethylation pathway of homocysteine). This methyl group is initially transferred to cob (I)alamin, to prevent methylcobalamin that is demethylated by homocysteine to form methionine by the cobalamin-dependent enzyme methionine synthase. [15] To maintain the enzyme activity, a reductive methylation by methionine synthase reductase is required. [16] Folate transport and metabolism are suggested to be under genetic control. [17] Folic acid and genetic maintenance Folic acid and its derivatives are important in several reactions in the single carbon transfer, which include the biosynthesis of purines, thymidines and the remethylation of homocystine to methionine. Normal folic acid and vitamin B12 metabolism were reported to be essential for normal development of the neural tube. [Figure - 2] shows the main steps of homocysteine and folate metabolism pathway. [18] Folic acid plays an important role in DNA metabolism; where it is required for the synthesis of dTMP from dUMP and the maintenance of DNA methylation. Under conditions of folic acid deficiency, dUMPs accumulate in the cell and get a better chance to be incorporated into the DNA instead of thymine. [19] There are good evidences suggesting that excessive incorporation of uracil in DNA not only leads to point mutations but may also lead to the generation of single- and double-stranded DNA damage, chromosome breakage, fragmentation and micronucleus formation. [20],[21] Folic acid and vitamin B12 are required for the synthesis of methionine and the common methyl donor S-adenosylmethionine (SAM), are required for the maintenance of methylation patterns in DNA that determine gene expression and DNA conformation. [22] When the concentration of vitamin B12 and methionine is low, SAM synthesis will be insufficient leading to the minimization of methylene tetrahydrofolate reductase (MTHFR) activity on the irreversible conversion of 5, 10-methylene tetrahydrofolate to 5- methyltetrahydrofolate, thus favoring an increase of uracil incorporation into the DNA. Deficiencies in folic acid and vitamin B12 therefore can lead to: (1) an elevated DNA damage rate and altered methylation of DNA, both are important risk factors in cancer [20],[21],[22],[23] and (2) an increased level in homocysteine status, an important risk factor for cardiovascular disease. [24],[25] The consequences of folic acid and vitamin B12 deficiencies may also play an important role in developmental and neurological abnormalities. [20],[21] Folate, homocysteine and neural tube defects It was reported that folate and folate derivatives are specifically linked to NTD in human since women who had pregnancies associated with fetal malformations had a higher incidence of aberrant folate metabolism [25] and a key role of folate in preventing NTD was suggested. [26] Maternal folic acid levels studies in the first trimester of pregnancy in mothers with NTD offspring tended to be lower than in the controls. Disorder of homocysteine metabolism was suggested to be the basis for NTDs [27] and was confirmed by the fact that homocysteine value in mothers with spina bifida affected offspring was 75% higher than the control. [28] Genetic basis of NTDs Many familial studies indicated a significant genetic component to NTDs (29). NTDs are associated with known genetic syndromes such as trisomies 21, 13 and 18, Meckel syndrome, anterior sacral meningocele stenosis and with various chromosome rearrangements. [30] Candidate genes studied in human NTDs Accordingly as it is articulated in the previous section, it is clear that folate deficiency is a contributor to the etiology of NTDs, and genes in this metabolic pathway have been the basis for many candidate gene studies. Several potential genes have been derived from folic acid pathway [Figure - 2]. [Table - 1] summarizes the results of studies on the NTD correlation with different genes in different populations. These genes included the polymorphisms of MTHFD1, MTHFR, MTR and others, including what population was studied. Conclusions and Recommendations The etiology of NTD is poorly understood, but it is now suggested that there is a complex interplay between environmental and genetic factors. [31] Knowledge of the association between NTD and various genetic markers related to development may help increase our understanding of the genetics of pathogenesis of NTD. [38] It is not easy to identify and determine the genetic risk factors of the NTDs because of the multiplicity of genes that participate in neurulation, and the importance of gene-environment interactions. Related researches of common genetic polymorphism especially that are enclosed in the folate one-carbon metabolism, indicate risk factors such as MTHFR, but no particular folate-related gene has yet been occupied as a major determinant of possibility for NTDs. Studies in a large-scale will be necessary to provide satisfactory statistical power to credibly test whether such polymorphisms are truly NTD susceptibility factors. [40] It will be more imperative too, if multiple genes (folate-related and others) were evaluated in the same persons in order to detect possible compounding belongings of combinations of risk alleles that, individually, might not be statistically significant. [17] References
Copyright 2011 - Indian Journal of Human Genetics The following images related to this document are available:Photo images[hg11031f1.jpg] [hg11031f2.jpg] [hg11031t1.jpg] |
| |||||||||


{kind=link}
{kind=link}
{kind=link}