 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Health, Population and Nutrition, Vol. 23, No. 1, March, 2005, pp. 44-51 Diversity of Vibrio cholerae Strains Isolated in Delhi, India, During 1992-2000 Shukla Das; Sunil Gupta; National Institute of Communicable
Diseases, 22 Sham Nath Marg, Delhi 110
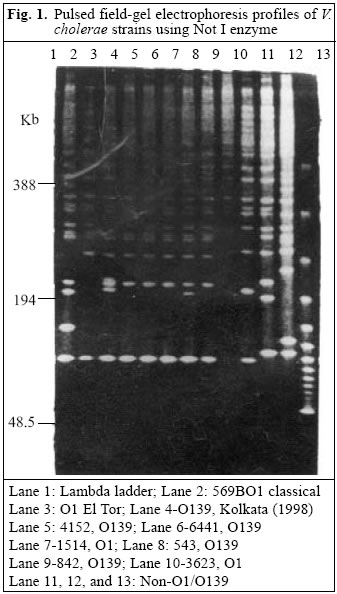
054, India Code Number: hn05006 ABSTRACT The National Institute of Communicable Diseases (NICD), Delhi, India, conducts a laboratory-based surveillance of cholera cases referred from the Infectious Disease Hospital, Delhi. The prevalence and antimicrobial susceptibilities of Vibrio cholerae O1, O139, and others, isolated from cholera patients for nine years, were analyzed to determine the changing trends in their isolation and drug-resistance patterns. In total, 29,196 stool samples or rectal swabs, collected during April 1992-December 2000, were included in this study. Of these, 13,730 (47%) were positive for V. cholerae: 11,091 for V. cholerae O1 (80.7%) and 1,943 (14%) for V. cholerae O139, and 696 (5%) were non-O1 and non-O139. V. cholerae O1 was the dominant serotype during 1992-1993, when V. cholerae O139 emerged as a new serotype but, thereafter, remained low from 1994 to 1999. Phenotypically, re-emerged V. cholerae O139 in 2000 displayed a difference compared to those that appeared in 1992-1993, in that the current O139 strains were sensitive to co-trimoxazole. Resistance to nalidixic acid and furazolidone was constantly high (100%) among strains of V. cholerae O1 and O139. All strains of V. cholerae were uniformly susceptible to chloramphenicol, tetracycline, amikacin, and norfloxacin. Molecular studies revealed different clones of V. cholerae O1 and O139 prevailing in the country with the re-emergence of V. cholerae O139 of a different clonality in Delhi in 2000, which is likely to play a critical role in temporal antigenic variation among the serogroups of V. cholerae. Key words: Vibrio cholerae; Cholera: Antigenic variation; Clonal diversity; Electrophoresis, Pulse-field gel; India Introduction Vibrio cholerae has been recognized as one of the common causes of bacterial diarrhoea throughout the developing world. Epidemics of cholera caused by toxigenic V. cholerae O1 and O139 (Bengal strain) represent a major public-health problem in most developing countries (1,2). Even non-O1 non-O139 serogroups of V. cholerae, though reported less frequently, are increasingly being shown as the causative agents of cholera (3). A dramatic shift in the aetiology of cholera was observed in the Indian subcontinent in the later part of 1992 and early 1993, when strains of a novel serogroup of V. cholerae O139 (Bengal strain) were associated with large outbreaks of cholera (4). In 1994, V. cholerae O1 El Torbiotype again replaced O139 as the dominant serogroup, causing cholera in Delhi (5). Following a latent period of three years, V. cholerae O139 surfaced again as an important pathogen of diarrhoea in 1998. In 2000, although V. cholerae O1 was once again the predominant pathogen, V. cholerae O139 also accounted for a large number of cholera cases and exhibited a different clonality, similar to V. cholerae O139 isolated in Kolkata. Recently, resistance to important antimicrobial agents has also been increasingly seen in the isolates of V. cholerae (6). Considering the shifting geographical and temporal patterns of V. cholerae O1 and O139, there is a continuing need to monitor the characteristics of strains and antibiotic-resistance patterns of this pathogen. The National Institute of Communicable Diseases (NICD), Delhi, is the nodal centre for carrying out national surveillance programmes for communicable diseases, of which surveillance for cholera is one of the major activities. This study reports the characteristics, including prevalence, antibiograms, and clonality of V. cholerae O1 and O139 recovered from stool samples of suspected cholera cases admitted to the Infectious Disease Hospital, Delhi. A comparative analysis of clonal relationships among toxigenic strains of V. cholerae was done using pulse-field gel electrophoresis (PFGE) at the Na-tional Institute of Cholera and Enteric Diseases (NICED), Kolkata. Materials and MethodsIn total, 29,196 stool samples or rectal swabs from patients with suspected cholera, admitted to the Infectious Disease Hospital during April 1992_December 2000, were processed in the Microbiology Division, NICD, using standard culture methods (7,8). Stool samples from each case were transported in Venkatraman Ramakrishnan (VR) fluid and rectal swabs in Cary-Blair medium and streaked on bile salt agar (BSA), thiosulphate citrate bile salt sucrose (TCBS), and McConkey agar and also inoculated into alkaline peptone water for enrichment and subsequent plating. Serogrouping was done by slide agglutination using polyvalent O1antisera CRI (Central Research Institute), Kasauli, and O139 antisera received from NICED. Antimicrobial susceptibility testing of representative strains of confirmed V. cholerae was performed on Muller-Hinton agar (DIFCO Labs, USA) employing the Kirby-Bauer disc-diffusion technique (9). Each isolate was tested against seven antimicrobial agents, e.g. tetracycline (30 mg), norfloxacin (30 mg), chloramphenicol (25 mg), furazolidone (50 mg), amikacin (30 mg), cotrimoxazole (25 mg), and nalidixic acid (30 mg). Phage typing of V. cholerae O1 and O139 was carried out at the Vibrio Phage Reference Laboratory, NICED, following the conventional Basu & Mukerjee methods and the new phage-typing scheme (10). Representative strains of V. cholerae O1 and O139, of each year, were subjected to polymerase chain reaction (PCR) using automated thermal cycler (Applied Biosystems, USA) with a gene Amp PCR reagent kit (Applied Biosystems, Roche, USA) for detection of ctxA and tcp El Tor genes, code for virulence factors, using the methodology of Saiki et al. (11) The ctxA, tcp El Tor primers were designed using an automated primer synthesizer in the Biotechnology Division, NICD. To ascertain the relationship and relatedness between the strains of V. cholerae, their restriction fragments were analyzed by cleavage of genomes using PFGE on a CHEF-mapper (Bio-Rad) at NICED. Details of the PCR are given below: The cycling conditions were 36 cycles of one minute at 94 °C, 1.5 minutes at 60 °C, and 1.5 minutes at 72 °C with a final extension at 72 °C for 10 minutes. A comparative analysis of clonal relationships among toxigenic strains of V. cholerae was determined using PFGE at NICED. Details of the PCR are given below: 
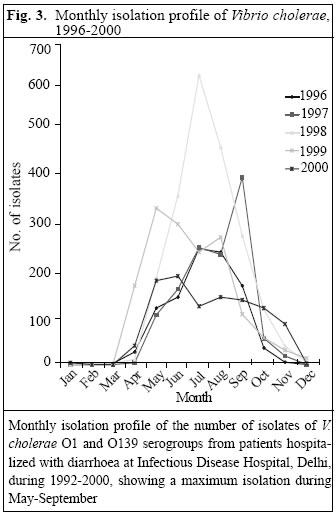
The cycling conditions were 36 cycles of one minute at 94 °C, 1.5 minutes at 60 °C, and 1.5 minutes at 72 °C with a final extension at 72 °C for 10 minutes. A comparative analysis of clonal relationships among toxigenic strains of V. cholerae was determined using PFGE at NICED. To perform PFGE, the genomic DNA of V. cholerae strains was prepared in agarose plugs (12). For complete digestion of the DNA, 50 U of Not I enzyme was used following the instructions of the manufacturers. PFGE of inserts was performed on a CHEF-mapper (Bio-Rad) with 0.5x TBE buffer (44.5 mM Tris, 44.5 mM boric acid, 1mM EDTA, pH 8.0). A DNA size l ladder was used as a molecular weight marker. Agarose gels were electro-phoresed at 14 °C using a constant voltage of 200 V at 120° with two different switch ramptimes. During the first run of 22 hours, the timings were 5 to 50 seconds to separate larger DNA fragments, and during the second run with 1.0 to 10 seconds to separate smaller DNA fragments for 12 hours. Gels were stained in distilled water containing 1.0 mg ethidium bromide per mL for 30 minutes, rinsed, and photographed. DNA was then transferred from gel to Hybond N+ membrane (Amersham International PLC, Buckinghamshire, England). Clonality of six strains of V. cholerae O1 (no. 1514 and 3623) and O139 (no. 4152, 6441, 543, and 842) was classified by determining their relatedness with standard reference strains of V. cholerae, i.e. classical O1 (569B), El Tor and O139, and Calcutta strain. Three strains of non-O1 non-O139 were also included (Fig. 1). Results During 1992-2000, a total of 29,196 stool samples of suspected cholera cases from the Infectious Disease Hospital were processed for enteropathogens. Of these, 13,730 (47%) specimens were positive for V. cholerae. Of the total cholera cases, the majority was V. cholerae O1 serotype, accounting for 11,091 (80.7%) isolates, followed by 1,943 (14%) O139 isolates; whereas 696 (5%) were non-serotypable. Figure 2 shows the year-wise distribution of V. cholerae during 1992-2000. In 1992, most V. cholerae isolates belonged to serotype O1. However, in 1993, a dramatic change occurred, with the emergence of a new V. cholerae serotype identified as O139 (Bengal strain) (13,14). Of 2,850 samples received during 1993, V. cholerae O1 accounted for 697 (45%) of the isolations, whereas V. cholerae O139 accounted for 831 (54.3%). In 1994, there was a re-emergence of V. cholerae O1 biotype El Tor, accounting for 1,754 (75.5%) isolates, predominating over V. cholerae O139 which accounted for 486 (20.9%) only. In subsequent years, from 1995 to 1997, the rate of isolation of V. cholerae O139 was much lower than that in any previous year. However, in 1998, V. cholerae O139 re-emerged, accounting for 11.1% of the isolations and, thereafter, following a quiescent period of one year, it again emerged in 2000, with isolations rising to 328 (31%). However, throughout this period, V. cholerae O1 biotype Ogawa remained the dominant serotype accounting for nearly 100% of the isolations, with scattered reports of Inaba serotypes (0.04-0.3%) seen during 1996-2000. Figure 3 shows the month-wise distribution of Vibrio cholera isolates from 1996 to 2000. As is evident, maximum isolations of cholera occurred during April-October, with a peak incidence observed during July-August. There were practically no isolations of cholera during the winter months of December-February, as seen in 1996-1998. The trend changed in the following year, where cholera cases continued to be reported, even during these months. Results of drug-sensitivity tests of randomly-selected strains of V. cholerae O1 and O139 from 1996 to 2000 are depicted in Table 1. The drugs depicting a high level of resistance to strains of V. cholerae O1 were nalidixic acid, furazolidone, and cotrimoxazole throughout the study period, whereas strains were usually susceptible to amikacin, tetracycline, norfloxacin, and chloramphenicol. Strains of V. cholerae O139 were susceptible to chloramphenicol, tetracycline, amikacin, and norfloxacin but sus-ceptibility to co-trimoxazole showed a changing pattern over the years, with 50% of strains showing resistance during 1996-997, which gradually became less resistant in the subsequent years. Strains were susceptible to nalidixic acid until 1997, but became increasingly resistance to the drug since 1998. The phage-typing pattern (Basu and Mukerjee scheme) of V. cholerae O1 strains showed a dominance of phage types 2 and 4 throughout the study period (1992-2000). However, when tested by the new typing scheme, since 1996, most strains were distributed among the phage type T27, the other less common phage types being 26, 3, 25, 19, 13, and 7 (15). With the recent development of a phage-typing scheme for V. cholerae O139, most of our O139 strains belonged to types 10 and 8 (Table 2) (16). The representative strains of V. cholerae O1 and O139 isolated during 1993-2000 were found to be positive for CtxA and tcpA (El Tor) genes by PCR. PFGE and genomic DNA restriction patterns after Not I restriction digestion of V. cholerae O1 and O139 strains, done at NICED, showed an interesting clonal relationship among the various strains (Table 3 and Fig. 1). Strains of V. cholerae O1 isolated in our laboratory in 1994 were closely related to V. cholerae O1 strains isolated in Kolkata during 1998, whereas V. cholerae O1 strains isolated in 1999 were similar to the strains isolated from the state of Kerala in 2000. Genetic analysis of V. cholerae O139 isolated in 1994 at NICD were similar to the Kolkata strains isolated during 1996-1997. How-ever, the genetic pattern of strains of V. cholerae O139 isolated during 1999-2000 at NICD were similar to strains isolated in Ahmedabad,north India, during an outbreak in 2000. Discussion Bacteriological and epidemiological investigations carried out in the cholera-endemic areas of Kolkata and Madras in earlier years revealed a dominance of V. cholerae O1 in acute diarrhoea cases (17-19). Reports of non-O1 vibrios causing diarrhoea gradually rose to 5-10% of all cases of cholera (20). It was late in 1992 when a new strain of V. cholerae O139 or Bengal emerged in Madras and spread to other Indian cities and other countries, e.g. Nepal and Pakistan (21). In this study, the distribution of cholera serotypes and phage types in the national capital territory of Delhi during 1993-2000 showed that V. cholerae O1 biotype El Tor serotype Ogawa was the predominant strain throughout the study period, in accordance with earlier reports (22,23) Most strains of V. cholerae O1 were clustered into phage type 2, following the Basu & Mukerjee scheme. However, from 1994 onwards, many earlier untypable strains analyzed by a new phage-typing scheme were found to be distributed among the new phage types T27 and T26, with lesser frequent types being T19, T13, and T7. Reports from other parts of the country yielded a wide variation in phage type distribution of V. cholerae O1, with the majority of strains clustered into types 17 and 18 (24). V. cholerae O139, after its first appearance in 1993-1994, remained latent for three years (1995-1997) and re-emerged in large numbers in 1998 and 2000. Such re-emergence at an interval of 1-2 year(s) is suggestive of its natural habitat being the aquatic environment remaining viable but in a non-culturable state. Therefore, constant surveillance of V. cholerae, including environmental surveillance, is necessary throughout the year (25). The antimicrobial susceptibility pattern of V. cholerae O1 showed a gradual change during the study period, with the majority of strains showing a high level of resistance to nalidixic acid, furazolidone, and co-trimoxazole. A similar trend has been reported elsewhere (26). Resistance to co-trimoxazole by V. cholerae O139 revealed a rapid shift from near 100% resistance in 1996-1997 to a relatively-susceptible pattern in 1998, probably due to its lesser use by clinicians. Also resistance to co-trimoxazole is encoded by a self-transmissible chromosomally-integrated transposon which, by virtue of its enhanced mobility, can lead to widespread dissemination of resistance and, on the contrary, its loss or absence can lead to enhanced susceptibility to the drug in V. cholerae strains (27). Based on the findings of this study, it is concluded that it is important to monitor the drug-resistance patterns of V. cholerae to prevent the emergence of multidrug resistance in the hospital setting. Based on the chronological evidence shown from this study, it appears that some drug-resistance expression might have transferred from one serogroup of V. cholerae to the other. The patterns of multiple antibiotic resistance determined in Delhi are comparable with those from other endemic locals in India and Bangladesh. Genetic studies revealed interesting analogies of V. cholerae isolated from different parts of the country. PFGE revealed V. cholerae O1 isolated in Delhi in 1994 to be similar to the Kolkata strain of V. cholerae O1 (iso-lated in 1998). V. cholerae O1 isolated in Delhi in 1999 was not only related to the Kolkata strains but also to V. cholerae O1 isolated from Kerala (South India), thus showing a widespread distribution of these strains in the country. V. cholerae O139 isolated at NICD, during 1994-1998, were genetically similar to strains isolated in Kolkata in 1994. However, V. cholerae O139 strains isolated in Delhi during 1999-2000 revealed genetic relatedness to Ahmedabad strains of V. cholerae O139 isolated in 2000, suggesting its circulation from west to northern parts of India. Based on the findings of this study, it is concluded that there is a constant change in the phenotypic and genotypic characteristics of V. cholerae, providing strong evidence for its environmental source and prevalent modes of transmission. It also indicates a continuous emergence of new clones of toxigenic V. cholerae, possibly through natural selection involving environmental factors and host immunity (28). AcknowledgementsThe authors are grateful to the National Institute of Cholera and Enteric Diseases, Kolkata, India, for assistance in carrying out the molecular analysis of Vibrio cholerae strains. Technical assistance of the staff of Cholera Laboratory, National Institute of Communicable Diseases, is also sincerely acknowledged. References
© 2005 ICDDR,B: Centre for Health and Population Research The following images related to this document are available:Photo images[hn05006t3.jpg] [hn05006f1.jpg] [hn05006f3.jpg] [hn05006t1.jpg] [hn05006f2.jpg] [hn05006t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}