 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Health, Population and Nutrition, Vol. 24, No. 1, March, 2006, pp. 1-5 Editorial Herd Protection and Herd Amplification in Cholera David A. Sack ICDDR,B: Centre for Health and Population Research, GPO Box 128, Dhaka
1000, Bangladesh
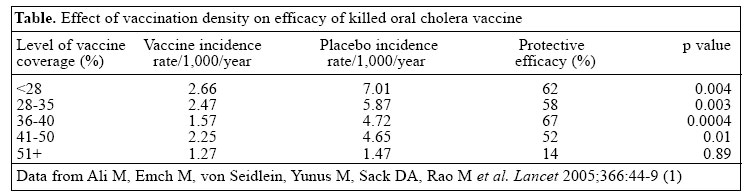
Email: dsack@icddrb.org Fax: +(880-2) 882 3116 Code Number: hn06001 Recent analysis of results of the 1985 vaccine trial in Bangladesh showed that a killed oral cholera vaccine could provide herd protection (1), and this finding sheds new light on the potential utility of this vaccine and other oral cholera vaccines. Although there may be several mechanisms for herd protection, this finding of herd protection was somewhat unexpected. The nature of the herd protection with cholera vaccine is unlike that with live oral polio vaccine which can be excreted and can infect others, thereby immunizing per-sons who did not receive vaccine directly. By contrast, the cholera vaccine used in this study was inactivated, making it impossible for non-immunized persons to be immunized inadvertently. Another type of herd protection is seen with vaccine for Haemophilus influenzae type b in which the vaccination reduces respiratory carriage of the pathogen, thereby eradicating the reservoir and reducing transmission. Another example is that of measles vaccination which essentially stops transmission when the density of susceptible subjects is reduced below that needed to sustain transmission. Since cholera is transmitted directly from contaminated food or water, the finding of herd immunity seemed not entirely expected. This editorial reviews the evidence for herd protection and introduces new findings from the environmental studies on cholera to suggest a more complete understanding of the mechanisms for herd protection with cholera vaccine. Hopefully, by combining the observations of ’herd protection’with some newer concepts of ’herd amplification’coming from recent environmental studies, we may develop a better understanding of the most efficient ways to control cholera. A review of the evidence for herd protection The first suggestion of herd protection came from one analysis of the 1985 trial in which young children who had not received vaccine were about 50% less likely to be infected if their mothers had received vaccine (2). It was hypothesized that mothers receiving vaccine would have increased levels of antibody in their breastmilk and that these antibodies might protect their young children. However, the antibody titres in breastmilk of vaccinated mothers were not different from that in mothers who were given placebo. The protection observed without a corresponding elevation in breastmilk antibody titres suggested a different mechanism not associated specifically with protection from breastmilk antibodies. It seemed more likely that the immunized mothers were less likely to be colonized with Vibrio cholerae and pass this organism on to their children. Thus, the children had some protection from the vaccine, although they themselves were not vaccinated. More direct evidence of herd protection came from the recent re-analysis of the same 1985 field trial which combined GIS analysis with the data of cases occurring during the first year of the trial (1). The re-analysis included 89,596 people who were individually randomized and were immunized with B-subunit-whole cell (BS-WC) vaccine, the whole cell (WC) only vaccine, or a placebo. In this trial, both BS-WC and WC only vaccines were protective and thus, for this analysis, the vaccinated groups were combined. This new analysis was intended to determine if there was evidence of an indirect protective effect among the 6,423 geographic clusters depending on the proportion of persons in the clusters who had received one of the active vaccines (either the BS-WC or the WC only vaccine). It was hypothesized that an indirect effect of the vaccine would be manifested by lower incidence rates of cholera among recipients of placebo in clusters with higher levels of vaccine coverage. Since the randomization was by individual, the proportion of persons receiving an active vaccine varied widely between the different clusters, with proportions receiving active vaccine between 4% and 65% in the different clusters. For this analysis, the unit of the cluster is the bari which is a group of houses within the village. Each bari is separated geographically from each other, thus forming small environmental microenvironments. As shown in the table, the incidence of cholera among recipients of placebo was inversely related to levels of vaccine coverage. The incidence among the recipients of placebo in the low-coverage clusters was about 7 per 1,000 during the first year. By contrast, the incidence among recipients of placebo in the highest-coverage clusters was only about 1.5 per 1,000; nearly a 5-fold difference. Also unexpectedly, the incidence of cholera among vaccinated individuals varied depending on the vaccine coverage from 2.7 to 1.3, a two-fold difference, suggesting that the risk of cholera among vaccinated persons was lower if most neighbours were also immunized. Thus, the analysis did establish an indirect protective effect among both immunized and non-immunized persons. An interesting feature of this analysis was the finding that the calculated protective efficacy was highly dependent on which control group was used in the calculation. Since the placebo clusters with high vaccine coverage had a lower incidence, there was minimal difference in rates between vaccine and control group in these clusters. A calculation based on a comparison of rates in placebo and vaccine recipients in these high-coverage clusters would have falsely concluded that the vaccine was not efficacious. On the other hand, if the incidence in recipients of placebo in the low-coverage clusters had been used for comparing with recipients of vaccine in the high-coverage clusters, the calculated protective efficacy would have been about 82%. It would seem that this estimate (82%) found in this analysis would simulate the protective efficacy which would have been ob-served if a cluster-randomization strategy had been used. It is possible that the field effectiveness might be even higher with an even higher-vaccine coverage. The limit of the indirect protective effect might not have been reached. A conclusion from this analysis is that, somewhat paradoxically, the observed protective efficacy was under-estimated in the 1985 trial because it used the ’gold-standard’, individually-randomized design, rather than being cluster-randomized. Some participants lived in clusters with a high proportion of individuals receiving the vaccine. Others lived in clusters with only a few receiving vaccine. Others lived in clusters with a gradient between the two extremes. Since the trial consisted of a mixture of baris with varying vaccine densities, it is highly likely that this confounding effectively reduced the observed efficacy during the 1985 trial. In the future, it would seem more appropriate to evaluate new cholera vaccines using a cluster-randomization procedure rather than individual randomization. Using this design, all consenting persons in the cluster should receive the same agent, either vaccine or control agent. A protection observed using this study design would provide information which includes the sum of the biological protection of the vaccine plus the herd protection afforded by the vaccination programme. The re-analysis from the 1985 trial clearly documented a substantial indirect protective effect from the oral cholera vaccine when a high proportion of the population was vaccinated. Thus, if vaccine programmes are planned, we can conclude that programmes will be more effective (and likely more cost-effective) if a high proportion of the population receives the vaccine. Efforts should be made not only to vaccinate susceptible persons, but extra effort should be made to achieve high rates of vaccine coverage during the programme. It was suggested that the explanation of the herd protection was related to the ’leaky’protection of cholera vaccine. That is, the vaccine does not provide absolute protection, but it does reduce the severity of the infection and lessens the colonization of bacteria. These two factors should lead to fewer numbers of organisms being excreted into the environment, and this likely leads to less transmission (3). The opposite of herd protection--herd amplification --and the role of the environment in herd protection If a cholera-vaccination programme is being planned, it is generally felt that the vaccine should begiven prior to the outbreak rather than waiting for the outbreak to begin. This seems logical from several perspectives. This strategy reduces the total numbers of persons becoming ill. It reduces the panic that sometimes accompanies an immunization programme during an epidemic. Studies have also found that this strategy is more cost-effective (4). The new understanding of vaccine-induced herd protection suggests that the vaccine may also be more effective if it can protect against transmission; thus, it seems logical to give the vaccine early. However, there may be additional reasons to consider vaccination prior to the outbreak or the season, and this relates to some new findings regarding the environmental reservoir, the initiation of the outbreak, and the ending of the outbreak. Environmental microbiologists and epidemiologists have attempted to explain the seasonality of cholera (5) and often focus on an attempt to predict when the cholera season will appear. They also have attempted to explain where the organisms go between seasons or outbreaks (6). More recently, efforts have been made to explain why epidemics seem to be self-limited (7). V. cholerae live in the environmental waters and co-exist with other members of the Vibrionaceae family and other waterborne bacteria. For reasons still not fully understood, but almost certainly related to climate and salinity, an environmental change increases the concentrations of these waterborne bacteria. However, there is no known mechanism for a selective increase in the pathogenic V. cholerae O1 or O139 in the environmental waters relative to other vibrios. The increase appears to be non-specific, and all the marine vibrios ’bloom.’ The pathogenic strains have no known virulence mechanism to ’out-compete’the other strains in this environmental bloom. However, during the bloom, it is thought that some persons become ’index infections’by drinking water (or water-contaminated food). After ingestion, the pathogenic V. cholerae (but not the other vibrios) multiplies in this person’s small intestine. Due to certain virulence factors of these pathogenic strains, including colonization factors, e.g. TCP and others, the pathogenic V. cholerae are selected for growth and are amplified. This index infection is probably asymptomatic, but the individual excretes the organism back into the environment in greater numbers than would result simply from the environmental bloom. These newly-excreted organisms, thus, are consumed in greater numbers by a second subject consuming the contaminated water or food. This second person with the infection then also excretes the organism, after intestinal selection and amplification, and the cycle between human and environment rapidly and selectively increases the number of the pathogenic strains in the environment. Not only are the pathogenic species amplified in terms of total numbers, these are also likely to be more virulent than the original infectious strain since the intestinal environment apparently upregulates virulence factors making these even more pathogenic. This increase in virulence properties, when passing through the intestine, has been seen when passing sequentially through a rabbit model (8), and upregulation of certain characteristics has also been seen in vibrios taken directly from human cases (9). Thus, it seems likely that persons living close to these ’index cases’are more likely to become infected. This initial cycling between the environment and humans also suggests a time lag between the climatic events which led to the first human infections and the actual start of the detectable outbreak by at least a few days to a few weeks. This new model suggests that the human intestine is a crucial link in the start of the epidemic. In fact, one should not conceptualize the habitat of vibrios as being environmental or human, but rather understand that the human intestine is part of the ecological niche of . As with other parasites, it may be possible to interfere with a part of the life-cycle and at least partially interrupt its transmission. While it seems logical that fewer persons, in general, who are excreting organisms into the environment would lessen the risk to others consuming water from this same environment, it also seems probable that the reduction in excretion will most effectively protect others if the vaccine were to protect against the infection of the index case within the cluster, thus, preventing the selection/ amplification which is responsible for the earliest phase of the outbreak. Ideally, one would like to prevent the index infections in persons who may be the key initiators of the outbreaks. Based on data from the 1985 trial, protection against severity was age-dependent, with younger children having more severe infection than older children and adults (10). This age-dependent effect might suggest that these non-immune younger subjects may be especially efficient as selection/amplifiers, and they might frequently harbour the index infection. Thus, immunization of children may be especially important if we wish to achieve herd protection even if the long-term protective efficacy may be less in this age-group. Alternatively, women who prepare food may be especially efficient in transmitting the vibrio to their families if they were first infected as was suggested in the 1985 trial (2). Historically, this concept of herd amplification makes sense. It has long been known that certain villages would be heavily affected, while other villages would be spared. Since these affected villages were often not contiguous, it seemed clear that this was not a matter of the strain spreading from place to place, but rather resulted from multiple infections which occurred simultaneously in different areas, but amplified locally in each individual area. Similarly, it has been noted that case to infection ratios are highly dependent on the rate of infection of the disease and the time during the epidemic. Although El Tor is often thought to have a lower case to infection ratio than Classical did, in fact, in Bangladesh at the height of the epidemic, the case infection ratio was similar in the two biotypes, being about 3 to 1. However, in other outbreaks when the rates of disease were lower, the ratio was much less, being about 40 to 1. This suggests that when many people are excreting the organisms, these organisms are more infectious, leading to herd amplification. New findings are also enlightening why cholera sea-sons end. Based on the work of Faruque et al., it is now apparent that some persons infected with V. cholerae excrete vibriophages, in addition to excreting V. cholerae (11). The persons infected early in the epidemic tend to be infected with bacteria, but not the phage. Later as the epidemic progresses, more phages are excreted and per-sons becoming infected later in the season consume both bacteria and phage. When ingested together, the phages are also able to multiply rapidly in the patient’s intestine since they exist in concert with their substrate, the vibrio. Thus, the patient is not only the amplifier of bacteria, but also the amplifier of the vibriophages. These phages are then excreted into the environment, and these attack and destroy the V. cholerae. It seems that the human hosts are both amplifiers of the epidemic and also amplifiers of the agent that leads to the collapse of the epidemic. We believe that this new knowledge may become useful in predicting and controlling epidemics in the future, but that the vaccines offer the best opportunity to control cholera at present. A final observation that may relate to the herd protection observed with the vaccine relates to the protection by sari filtration of water. With the hypothesis that cholera vibrios are associated with copepods and that these copepods can be filtered using sari material, an intervention was developed in which residents of 65 villages in Matlab, using a village-randomized design. They were taught and encouraged to filter their household water. The intervention villages experienced a 48% reduction in cholera compared to the control villages (12). Is it possible that this protection was most efficient in protecting against the index infections and, thus, the field effectiveness was much greater than its biological power? Limitations of herd protection The fact that herd protection can occur seems well-established by the recent analysis of the 1985 vaccine trial in Matlab; however, there may be limitations to the applicability of these findings to all situations where cholera vaccine may be used. The Matlab environment is typical of the rural areas of South Asia where cholera is endemic. Most people live in a bari, or cluster of houses that are separated geographically from each other. Usually, the bari has its own water supply. Several baris constitute a village. This is quite different, however, from the urban areas where persons live in densely-populated neighbourhoods, share water sources with many other people, and often have sanitary facilities that are shared by many other families. When floods occur, water and sewage become mixed, and cholera epidemics have resulted from the grossly-contaminated environment (13). In the urban environment, it, thus, seems doubtful that herd protection would alter the course of the epidemic and protect non-immunized persons. Whether herd protection would be active in other situations where cholera epidemics are a risk, such as refugee camps, is unknown. These situations may be intermediate between the rural and the urban environment. The field effectiveness of the vaccine programme would depend on whether the vaccine could prevent the index cases from amplifying the pathogen within the camp and the extent to which the excreta from a few index cases would contaminate the general environment, especially the water supply. Implications There are several implications of these new observations on the control of cholera. First, if one intends to immunize populations for cholera, it will be important to achieve a high rate of immunization coverage to provide a higher degree of field effectiveness than would occur if the coverage were low. Second, the vaccine should be given prior to the epidemic in an attempt to prevent the first index cases; thus, methods to predict epidemics will be especially important. Third, if there were identifiable groups who play key roles in the early stages of herd amplification, e.g. mothers or small children, these groups should especially be targeted for vaccination. Finally, the concepts of herd amplification suggest that vaccines and improved water and sanitation should protect synergistically. By breaking the transmission cycle between humans and the environment through improved water and improved immunity, it may be possible to greatly reduce incidence. Thus, vaccines and sanitation should not be viewed as competing strategies, but rather part of the same strategy to prevent transmission. REFERENCES
© 2006 ICDDR,B: Centre for Health and Population Research The following images related to this document are available:Photo images[hn06001t1.jpg] |
| |||||||||

{kind=link}