 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
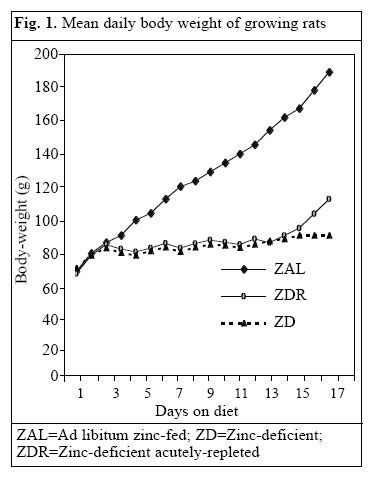
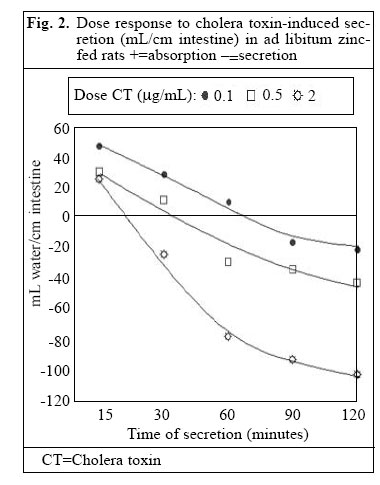
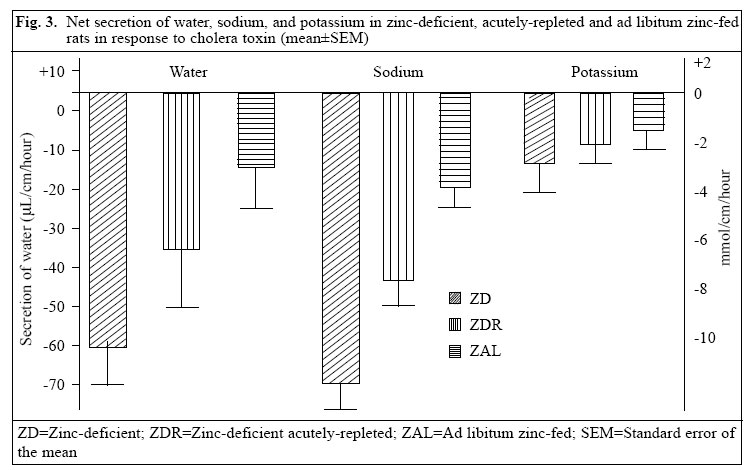
Journal of Health, Population and Nutrition, Vol. 24, No. 1, March, 2006, pp. 42-47 Impact of Zinc Deficiency on Vibrio cholerae Enterotoxin-stimulated Water and Electrolyte Transport in Animal Model S.K. Roy1, Andrew M. Tomkins2, Gulshan Ara1, S.P. Jolly1, Wajiha Khatun1, R. Chowdhury, and Barnali Chakrabarty1 1Clinical Sciences Division, ICDDR,B: Centre for Health and Population Research, GPO Box 128, Dhaka 1000, Bangladesh and 2Centre for International Child Health, Institute of Child Health, 30 Guilford Street, London WC1N, 1EH, UK Correspondence and reprint requests should be addressed to: Dr. S.K. Roy, Senior Scientist, Clinical Sciences Division , ICDDR,B: Centre for Health and Population Research, GPO Box 128, Dhaka 1000, Bangladesh, Email: skroy@icddrb.org, Fax: 880-2-8823116 Code Number: hn06007 Abstract The effect of zinc deficiency on the function of the intestine to absorb water and electrolytes was studied in animal models, stimulated by Vibrio cholerae enterotoxin. Sprague-Dawley rats, used in the study, were divided into four groups: Zinc-deficient, ad libitum zinc-fed control, zinc weight-matched control, and zinc-deficient acutely-repleted. 14C-labelled polyethylene glycol solution was used for measuring the absorption capacity of the small intestine. Significantly lower absorption of water and sodium per cm of the intestine was observed in the zinc-deficient animals compared to the ad libitum zinc-fed control animals (p<0.01). An improved absorption capacity was equally observed in the zinc-deficient acutely-repleted animals and ad libitum zinc-fed control group. The zinc-deficient animals showed four times greater cholera toxin-induced net secretions of water and sodium compared to the ad libitum zinc-fed group (p<0.01), while a 40% reduction was observed in the zinc-deficient acutely-repleted group. The results suggest that zinc deficiency is associated with reduced absorption of water and electrolytes and increased secretion of the same stimulated by cholera toxin. Key words: Zinc deficiency; Cholera toxin; Water-electrolyte balance; Intestinal secretions; Perfusion; Disease models, Animal Introduction The clinical genetic syndrome of zinc deficiency-- acrodermatitis enteropathica--characterized by skin lesions and chronic diarrhoea, improves rapidly following supplementation with zinc (1). Zinc deficiency in malnourished children with simultaneous diarrhoea has also been reported (2). Supplementation of zinc showed a 28% reduction in stool output, 14% reduced duration of acute diarrhoea (3), and 33% reduction in the recovery period of persistent diarrhoeal episodes among under-weight children (4). Results of pooled analysis of studies on supplementation of zinc conducted in different coun-tries have further confirmed the beneficial effect of zinc in acute and persistent diarrhoea (5). Results of a study on guinea pigs have shown that malnutrition is associated with increased intestinal permeability to small molecules and that pharmacological doses of zinc prevent such abnormality (6). Zinc improved intestinal permeability in children with acute and persistent diarrhoea in Bangladesh (7). Experimental zinc deficiency is associated with reduced enzymatic functions of the small intestine and ultrastructural abnormality of the intestinal epithelium and paneth cells in zinc-deficient rats (8). The permeability of the mucosal cell membrane may also be altered in zinc deficiency, and stabilization of the cell membrane is affected in zinc deficiency leading to defective transport of sodium (9,10). It has been shown that net transport of water and sodium from the small and large intestines of zinc-deficient rats was significantly decreased compared to the corresponding mean values for pair-fed controls and for ad libitum zinc-fed animals (11). On the other hand, supplementation of zinc increased transport of sodium in leukocytes of previously-malnourished children during nutritional rehabilitation (12). Furthermore, sodium flux was improved in renal tubular cells and erythrocytes after supplementation of zinc (13,14). Since there is no information on mechanism of reduction of diarrhoea by supplementation of zinc, these observations led us to investigate the role of zinc on the transport of water and electrolytes in response to V. cholerae enterotoxin using perfusion studies in animal models. Materials and Methods Study site The study was conducted at the Nutrition Research Unit of the Department of Human Nutrition, London School of Hygiene & Tropical Medicine (LSHTM), UK. Study animals Three-week old Sprague-Dawley rats weighing about 70 g each were divided into four groups. The animals were housed in individual stainless steel cages in a room maintained at 24-25 oC with an alternating cycle of l2 hours of light (8:00-20:00 hours) and l2 hours of dark. Animals were fed and weighed daily between 10:00 and 11:00 am using a Salter Balance (SD l000 T/0.0l CMS Weighing Equipment Ltd., Middlesex, UK). The zinc-deficient animals were fed a semi-synthetic diet containing 0.8 μg/g of zinc for l6 days. The ad libitum zinc-fed control animals were fed a similar diet containing 55.0 μg/g of zinc for the same period. A group of zinc-deficient animals was acutely repleted by feeding the zinc-containing diet (55.0 µg zinc per g) ad libitum for 48 hours. The other group of animals (zinc weight-matched controls) was fed the zinc-containing diet (55 μg/g) until their body-weight was equal to that of zinc-deficient rats. This group was included to con-trol the effect of age and zinc status in respect to absorptive function. Food intake was measured at every 24 hours using the Salter Balance (Salter UK) with sensitivity of 1 mg. The composition of diet is shown in Table 1. Perfusion technique The perfusion system was similar to the method described by Sladen and Harris (15). Before perfusion, rats were fasted for l2-l8 hours, except for the allowance of distilled de-ionized water ad libitum. Anaesthesia was maintained by subcutaneous pentobarbitone (5.4 mg/l00 g of body-weight), while the body temperature of the animals was maintained at 37 oC with the help of an angle point light. Rectal temperature was monitored with a digital thermometer. The abdomen was opened by a midline incision, and the proximal part of the jejunum was cannulated l0 cm distal to the ligament of Treitz through a small transverse slit. A polyethylene cannula of l-mm diameter was secured with silk at the proximal entry. A segment of 25-30 cm from the proximal entry was obtained, and a similar cannula was secured at the distal end. The segment was replaced into the abdominal cavity and was covered with a piece of surgical gauze soaked in physiological saline. The vascular supply of the intestine was kept intact. The inlet cannula was connected to tubing through a peristaltic pump (Minipulse 2, Gilson Medical Instruments, France) to maintain per-fusion at a constant rate of 0.44 mL per minute. A closed mouth flask containing perfusion fluid was kept in a water bath at 37 oC. Basic perfusion solution in the flask contained electrolytes and marker with (mmol/L) NaCl l30, KCl 4, glucose 2, NaHCO2 25, 14C polyethylene glycol 5 μCi, and unlabelled polyethylene glycol 4000 3 g with a pH 7.0. After an initial equilibrium period of 25 minutes, serial collections of perfusate for each l5 minutes were obtained from this one-way single-pass perfusion. Net transport of water and electrolytes was estimated by measuring the change in concentration of 14C polyethylene glycol. Calculation of water or electrolyte transport The following formula was used for calculating water and solute transport:
A negative sign indicates net secretion, and a positive sign denotes net absorption. i=signifies inlet fluid, o=outlet fluid, v=volume perfused (rate x period), S=solute, and DPM=discharge per minute of isotope. Data analysis Concentrations of sodium and potassium were deter-mined with an automatic analyzer (KNA l, Radiometer, Copenhagen). An average hourly rate for absorption or secretion was calculated by taking the average value of at least three samples. The perfused segment of the intestine was stripped out of the mesentery and was then gently pressed between the fingertips to remove water from the lumen. The length of the segment was measured using a vertical scale with a constant hanging weight of 5 g and weighed in a balance. Mucosa from non-perfused segments of the intestine was also scraped with a microscopic slide, and mucosal weight per cm of the intestine was calculated. A test of statistical significance was performed by unpaired Student’s t-test, and a p value of less than 0.05 was considered significant. Ethics No human subjects were used in the study. The animal experimentation committee of LSHTM approved the protocol. Results Growth of experiment animals Significantly greater weight gain was observed in the ad libitum zinc-fed control animals compared to the zinc-deficient animals. Figure 1 shows the mean (±SD) daily body-weight of rats on zinc-deficient and zinc-containing diet. The final mean body-weight of the zinc-deficient animals after l6 days of zinc-deficient diet reached from 70 g to 90 g compared to 180 g of ad libitum zinc-fed control animals (p<0.01). After 48 hours of zinc repletion, the zinc-deficient rates increased their body-weight by 24% to a mean of ll0 g (zinc-deficient acutely-repleted). The zinc weight-matched controls reached the body-weights similar to the zinc-deficient group but were younger in age. Transport of water and electrolytes without stimulus of cholera toxin A marked difference in net transport of water and electrolytes per unit length of intestine was observed among animals of different zinc status (Table 2). The zinc-deficient animals absorbed only 43% of that absorbed by the ad libitum zinc-fed control group (p<0.0l). In the zinc-deficient acutely-repleted animals, net absorption of water increased nearly equal to that of the ad libitum zinc-fed control group (p>0.0l). Sodium trans-port followed a similar pattern of water absorption. The zinc-deficient animals absorbed less than 50% of the sodium absorbed by the ad libitum zinc-fed control animals (p<0.05). Forty-eight-hour zinc supplementation resulted in an improvement of sodium absorption of the zinc-deficient group equal to that of the ad libitum zinc-fed control animals. No difference in absorption of potassium was observed between any pair of experimental groups. The weight-matched controls had similar absorption as the zinc-deficient group. Dose response of V. cholerae enterotoxin Figure 2 shows that a higher dose of V. cholerae entero-toxin in the perfusion fluid induced a higher amount of water secretion in the ad libitum zinc-fed control animals. The initial net absorption varied inversely with the con-centration of enterotoxin. The onset of net secretion was 22 minutes after perfusion of 2.0 μg CT/mL, after 55 minutes with 0.5 μg CT/mL, and after 85 minutes with 0.l μg CT/mL. The severity of secretion varied directly with increase in toxin concentration. Net secretions at two hours were l8 mL, 50 μL, and ll0 μL/cm with 0.l μg, 0.5 μg, and 2.0 μg CT/mL respectively. Challenge with V. cholerae enterotoxin The smallest dose was chosen to produce a modest secretion to enable the detection of the additional effect of zinc nutritional status of the models. V. cholerae entero-toxin was added in the perfusate at a concentration of 0.l μg/mL. Net transport of water and electrolytes was observed as net secretion (Fig. 3). The zinc-deficient group showed nearly four times higher secretion of water than that exhibited by the ad libitum zinc-fed controls. The zinc-deficient acutely-repleted animals demonstrated a 40% reduction in secretion of water compared to the zinc-deficient group. Net secretion of sodium was ob-served in all the groups. The zinc-deficient group had almost four times greater sodium secretion than that of the ad libitum zinc-fed control group, while approximately 60% reduction was observed in the zinc-deficient acutely-repleted group. Secretion of potassium increased about two folds during zinc deficiency, which reduced to 30% after 48 hours of supplementation with dietary zinc. DiscussionThe role of zinc in the absorptive and secretary function of intestinal mucosa was examined in this study. It was evident that the net absorption of water or sodium was reduced in the zinc-deficient animals compared to the ad libitum zinc-fed control or repleted animals. Repletion of zinc through the diet for only 48 hours improved absorption of water and sodium compared to the zinc adequate animals with adequate zinc status. It is likely that the active site of absorption was influenced by supplementation of zinc in these animals. Growth of mucosal tissue might be enhanced after supplementation of zinc (16), which could be associated with the improvement in transport. As most absorptive cells are villous tip cells, it is possible that the improvement in net absorption is due to improvement of this cell population. Improvement of mucosal integrity (17), permeability (12), or enzymatic activity (18) in absorptive cells might have helped further net absorption. The zinc weight-matched controls were younger than the zinc-deficient group, yet the absorptive capacity in this group looked better than that of the deficient group, although statistical significance was not reached. This implies that, although mucosal mass per-unit length in the zinc weight-matched controls is less compared to the zinc-deficient group (10.1 vs 13.3 mg/cm), the absorptive capacity was not less in the zinc weight-matched animals than in the zinc-deficient adult animals. A plausible explanation might be that mucosal functions in the zinc weight-matched group were not compromised, as they were not zinc-deficient. Zinc deficiency might result in damage of the epithelial cells (19) and mucosal lining of the gastrointestinal and pulmonary tracts (17). It is known that sodium transport is associated with coupled active absorption of sugar, amino acids, or urea (20). As the glucose concentration of the perfusion solution in this study was only 2 mmol/L, coupled carrier transport of electrolytes and water was not responsible in this perfusion model. In response to the secretory stimulus of cholera toxin, the zinc-deficient animals secreted more water, sodium, and potassium compared to the ad libitum zinc-fed controls. Net secretion of water and sodium was reduced after 48 hours of acute zinc repletion to a comparable level of the ad libitum zinc-fed animals. The improvement in absorption could be related to the regeneration of mucosal tissue (21) as a result of zinc repletion even if it was for only 48 hours. The mechanism of cholera toxin-induced secretion involves activated adenylate cyclase, which increases cyclic AMP concentration in the epithelial cell resulting in active secretion of water and electrolytes. Malnourished rats are more susceptible to epithelial cyclic AMP accumulation and increased secretions of sodium and potassium in luminal fluid when perfused with lactose solution (22). We did not, however, measure the cyclic AMP in the mucosa. The study provides the basis to consider that better zinc status may lead to better mucosal structure and function to improve transport of water and electrolytes. Dose response of V. cholerae enterotoxin showed clear effects of dose on the timing of onset of secretion. In respect to initiation of secretion, higher to lower concentration gradually delayed the time of onset of secretion. Compared to other perfusion studies with cholera toxin (23,24), our model was consistent in time lag for initiation of secretion. The study demonstrated the associations among zinc deficiency, growth retardation, and effect of supplementation of dietary zinc on body-weight of the experiment animals. Despite very low zinc in the diet, the animals increased in body-weight over the first three days, then there was very little growth from the fourth to seventh day, and thereafter no weight gain was observed over the rest of the two-week period. Slow growth from the third day and afterwards indicated zinc deficiency as a growth-limiting factor. Growth retardation in severe zinc deficiency observed in our study was comparable to other studies (25,26). The zinc nutritional status was related to intestinal mucosal integrity and absorption of water and electrolytes in this study. Supplementation of zinc in diet also reduced the net secretion of water and electrolytes induced by cholera toxin. This may explain the role of zinc in reducing diarrhoea in children. AcknowledgementsThe study was carried out with the fellowship grant from the United Nations University (first author) and was con-ducted at the Nutrition Research Unit of the Department of Human Nutrition, London School of Hygiene & Tropical Medicine, UK. Partial financial support was given by ICDDR,B: Centre for Health and Population Research. ICDDR,B is supported by countries and agencies that share its concern for the health and population problems of developing countries. The authors acknowledge with gratitude the commitment of donors to the Centre's research efforts. References
© 2006 ICDDR,B: Centre for Health and Population Research The following images related to this document are available:Photo images[hn06007t1.jpg] [hn06007f1.jpg] [hn06007f3.jpg] [hn06007t2.jpg] [hn06007f2.jpg] |
| |||||||||

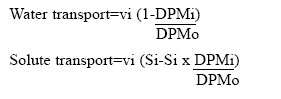
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}