 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Health Sciences, Vol. 2, No. 1, April, 2002, pp. 2-10 ORIGINAL ARTICLES Screening of extracts of Zanthoxylum chalybeum and Warburgia ugandensis for activity against measles virus (Swartz and Edmonston strains) in vitro *Olilaa D, Olwa-Odyekb and Opuda-AsibocJ. Departments of Veterinary Physiological Sciencesa,
Pharmacyb and Public Health and Preventive
Medicinec,Makerere University, Box 7062, Kampala, Uganda. Code Number: hs02029
ABSTRACT
A large proportion of the population in Uganda still relies on the use of plant extracts for treatment of various ailments. This study tested the claimed efficacy of some plants in the treatment of measles. In vitro antiviral assays were performed on extracts of two medicinal plants (Warburgia ugandensis and Zanthoxylum chalybeum) using measles virus (Edmonston and Swartz strains) as the test organisms. The assays performed were the neutralisation tests and the plaque reduction assays. Of the two plants Z. chalybeum had demonstrable in vitro antiviral activity in the seed extracts (titer reduction factor [TRF]: 100, for the ethanolic extract). The in vitro antiviral activity of the seed extracts was demonstrated to be due to compound 27-135D (TRF=1000), which was characterized by 1H-NMR spectroscopy as the alkaloid skimmianine. Skimmianine had minimal toxicity to VERO cell lines. The petroleum ether extracts and the ethanolic extracts of Warburgia ugandensis had no inhibitory effect on cytopathic effect (CPE) formation, especially at the maximal non-toxic dose (MNTD). The extracts of W. ugandensis were highy toxic to VERO cell lines. The TRF values for the stem bark extracts of W. ugandensis were: water extract, 10; ethanolic extract, 1; fraction 27-163D, 100., which were regarded to be too low. Seed extracts of Z.chalybeum therefore probably cure measles due to the antiviral effect of skimmianine. It is not clear how extracts of W. ugandensis produce a beneficial response in measles disease, if at all. INTRODUCTION Many people in Uganda live in rural areas, several kilometers from health and veterinary centers. Because of this, a large proportion of the population continues to rely on traditional methods of treatment, using herbal extracts, which have been claimed to produce a beneficial response. While modern human and veterinary medicine have gained a great deal of credit in both the curative and preventive aspects of disease control, there still remains a lot that is unknown or is unsatisfactorily known and therefore more research needs to be done. For instance in the field of chemotherapy, several diseases including tuberculosis and brucellosis still require very protracted treatment in order to achieve a cure. There are some diseases, which in the past were easily curable using drugs, but are today very difficult to successfully treat as a result of drug resistance by the etiologic agent. There are also some diseases for which cure is speculative or unknown. The present modern system of treatment (both -human and veterinary) is inaccessible to many and expensive. It has therefore become necessary to study ways of improving an already available medical service which uses medicinal plants. Past experience has shown that many useful drugs can be derived from plants (eg. atropine and ergometrine). Measles is one of the main killer diseases of children in Uganda. There is currently no treatment for the primary viral disease. Most treatment is directed at using antimicrobials to control secondary infections. In Uganda, plants may represent a potential source of drugs against measles. Two medicinal plants (Z. chalybeum and Warburgia ugandensis) have been reported to be of value in treating measles. The use of such plants could be attributable to their antimicrobial activity (against secondary bacterial infections) or to their antiviral action against the measles virus. Here we report the results of studies that were designed with the objective of screening extracts of the two plants against the measles virus in vitro, in order to shed some light on their potential value as sources of drugs. MATERIALS AND METHODS
i) Plant collection and pre-extraction preparation
Zanthoxylum chalybeum specimens were collected from Ongino, Kumi district, Eastern Uganda. Botanical identification was done by a botanist at the Natural chemotherapeutic Research Laboratories (Wandegeya), Ministry of Health, Kampala. Voucher specimens have been kept at the Department of Pharmacy, Makerere University Medical school, for reference purposes. The stem bark was chopped into small pieces and dried under the sun for three weeks. The seeds were already dry when collected, but they were also further sun-dried for one week. Warburgia ugandensis stem bark was collected from Mabira Forest, 20 km on Kampala-Jinja road and also botanically identified and voucher specimens kept as given above. The stem bark was likewise chopped and sun dried for three weeks. The dried specimens were then crushed by pounding in a wooden mortar into fine powder, ready for extraction. The powdered materials were macerated in petroleum ether and alcohol as solvents. The water extracts were obtained by soaking in water.
ii) Plant extraction, purification and spectroscopic procedures
One hundred grams of powdered plant materials of Z. chalybeum and W. ugandensis was soaked in petroleum ether, ethanol and water respectively in Ehlenmeyer flasks, corked and allowed to stay overnight. The marcerate was filtered with Whatman No. 1 filter paper. The filtrate (ethanol and pet ether) was concentrated in vacuo in a roundbottomed flask using a rotary evaporator (Heidolph model) under low temperature. The water extract (after filtration) was poured onto watch glasses and petridishes and dried in air in a hood. The extract was then scrapped off onto vials-ready for the phytochemical analysis and the bioassays. Melting points were determined on Koffler hot stage apparatus and on a Thomas Hoover capillary melting point apparatus. Infra-Red spectra were run using KBr discs on Perkin Elmer Infrared spectrophotometer, model 727B. 1H NMR spectra were determined for solutions in deuteriochloroform and DO with a 2 MHz, tetramethylsilane as internal standard. Analytical thin layer chromatograms were run on 0.2mm thick layer of silica gel, (Merck). The products isolated were detected by UV fluorescence. Preparative layer chromatography (PTLC) was run on 1.0mm thick layer of silica gel. Column chromatography was performed on silica gel 60 (70230 mesh) and Sephadex LH-20. a) Extraction and isolation of compounds from Zanthoxylum chalybeum Crushed seeds of Z. chalybeum (100g) were soaked in ethanol, placed in a shaker for 24 hr, filtered, and concentrated under reduced pressure, to obtain 6.9g of the crude extract. Compound 27-135D crystallized out of the crude extract and was filtered off and re-crystallized from methanol. Stem bark of the plant (100gms) was also soaked in ethanol, placed in shaker overnight, filtered and concentrated under reduced pressure to obtain 7.6g of the crude extract. The crude extract was chromatographed over silica gel column (flash) and eluted with Pet/EtOAC and then EtoAC:MeOH mixtures of increasing polarity to obtain a total of 28 fractions. Fractions 21 and 22 (Solvent systems EtoAC:MeOH (6:4) and EtOAc:MeOH (1:1)) yielding a white crystalline compound 27-167A (20mg). Fraction 1, solvent system: EtOAc:MeOH (8:2) yielded an alkaloid, 27-165A. Fractions 2-6, solvent system: EtOAc:MeOH (1:1) yielded alkaloids 27-173A and 27173B after purification using PTLC using chloroform alone as the solvent system. b) Extraction and isolation of compounds from W. ugandensis. The stem bark (100g) of W. ugandensis was soaked in EtOH, placed on a shaker for 24 hours, filtered, concentrated to obtain the crude extract (13.2g). The crude extract (6.6g) was dissolved in 80% aqueous MeOH (100ml) and extracted with petrol (100ml x 3). The petrol extract was dried over anhydrous Na2SO4., filtered, concentrated under reduced pressure to obtain 0.48g of extract (27-163A). To the aqueous layer, distilled water (33.3ml) was added and then extracted with chloroform (2 x 100ml). The chloroform extract was dried over anhydrous Na2SO4., filtered, concentrated to obtain 1.42g of extract (27-163B). The remaining aqueous extract was dried in the hood to obtain 2.40g of 27-163D. Fractions 27-163A which showed highest activity was applied on Sephadex LH-20, eluted with hexane-CH2Cl2 (1:4) and then hexane-acetone (3:2) to obtain eight fractions. The first and the last fractions were blank on analytical TLC and were hence discarded. The second fraction (27-169A) was further purified on PTLC, benzene:petrol:EtOAc (3:9:1) and benzene:petrol:EtOAc (3:4:3) to obtain, respectively, compounds 27-179J, 27-179K, 27-179L, 27-179M an 27-179A. Furthermore, ground stem bark (200g) was macerated first with petrol and then with Ethyl acetate (EtOAC). The petrol extract was concentrated to yield 10g of crude extract, which was fractionated over silica gel column eluting with petrol and then petrol containing increasing amounts of EtOAC. In order of elution, the following fractions were obtained: 49-169b (1.17g), 49169c (500 mg), 49-169d (500mg), 49-169e (1.15g), 49169f (1.15g), 49-169g (1.16g), 49-169h (150mg), 49-169i (651mg), 49-169j (1.56mg), 49-169k (850mg), 49-181a (665mg). Fraction 49-169k was pure muzigadial.
iii) In vitro activity against measles virus a) Virus type and
source
The measles virus used for the neutralization test (cytopathic effect (CPE) inhibition) was the lyophilized suspension of attenuated Schwartz strain grown in chicken embryo cells (MorbilvaxR). Each 0.5ml dose contained 10,000 TCID50 of virus and 50mcg each of neomycin B sulfate and kanamycin. It was obtained from the National Public Health Laboratories, Nairobi. The virus used for the plaque reduction assay was the Edmonston strain measles virus (Moraten Berna®). The World Health Organization supplied it. It was designated to contain 4.0log PFU’S per 0.5ml when reconstituted
b) Effect of extracts on monolayers (maximal nontoxic dose)
100mg/ml of each of the extracts (crude and the fractions) were made using dimethlysulfoxide (DMSO) as solvent. 0.5ml of this stock solution was added to 4.5ml of maintenance medium containing antibiotics. Serial twofold dilutions of the extracts were done in bijou bottles using M199 as diluent. One drop (using a standard microtiter dispenser) of each dilution, corresponding to 50ul of extract, was dispensed in triplicate to microtiter plates previously prepared (i.e. with monolayers of vero cells). The plates were incubated at 370c in a carbon dioxide incubator and the cells observed for 5-7 days for any changes attributable to the extract. Some of the wells were treated with dimethyl sulfoxide alone. Serial two-fold dilutions of measles antiserum were also dispensed in triplicate. The results were scored after 7 days of incubation at 37oC in a carbon dioxide incubator.
c) Determination of virus infectivity titers
Maintenance medium (M 199) was used as diluent to reconstitute the viruses. 5ml of M 199 was introduced per vial, mixed to give the seed viral suspension. 0.9ml of M 199 was further mixed with 0.1m1 of the seed viral suspension and serial ten fold dilutions made. One drop of the viral preparation above (corresponding to 50ul of virus) was applied on micro titer plate in quadruplicate per dilution onto confluent VERO cells and incubated at 370c in a carbon dioxide incubator. Observation of cytopathic effect (syncytia formation) was made over a 5-7 day period during which syncytia formation was evident. The wells that served as cell controls did not receive any virus challenge. The calculation of the TCID50 was based on the Reed and Muench formula (1).
d) Neutralization test (inhibition of cytopathic effect (CPE) Constant
virus-varying extract method
Here a constant dose of the virus (Morbilvax®) containing 100 TCID50 was used based on the titration procedures already described. Serial two-fold dilutions of extracts in M 199 starting from the MNTD of extract was made. Each MNTD of extract was designated as the “neat”and serial dilutions made. The virus was also diluted using M199 to obtain 100 TCID50 per drop of 50ìl (using a standard micro titer dispenser). For every dilution of extract and serum, one drop of extract (or serum) was reacted with one drop of virus in a “reaction” plate without cells. The mixture was gently agitated to ensure complete mixing of the extract-virus mixtures. The lid was replaced and the reactants left at room temperature for one hour. After one hour the extract (or serum) - virus mixtures were transferred to monolayer cultures (VERO cell line) grown on micro titer plates, using a separate pipette for each inoculation. The inoculated cultures were incubated at 370c in a carbon dioxide incubator and observed daily for CPE. The un-inoculated wells served as cell control while the wells with extract alone or serum alone served as extract and serum controls respectively. Some wells also received 50ul of virus alone - being the virus controls.
Constant extract-varying virus method
Here a constant concentration of extract or serum previously determined to cause inhibition of CPE was used. Tenfold dilutions of measles virus were made in M 199. Equal volumes of virus (one drop) and extract were reacted on a plate for one hour and then later transferred to the cell cultures.
End point titration technique
Equal volumes of extract and virus were reacted for 30 minutes (0.5ml virus + 0.5ml extract at MNTD). Serial tenfold dilutions of the reactants (in M199) were made in 48 well-plates. The 96-well microtitre plates, previously monolayered with vero cells as given in 3.6.1, were preemptied to give room for the reactants.Two drops (50µl each) of the serially diluted extract and virus mixtures were dispensed onto the 96-well plate using standard microtitre dispensers. The reactants were incubated at 37oC for 5-7 days in a carbon dioxide incubator and observations made for syncytia formation. Antimeasles serum was used as a positive control while virus alone plus M199 served as virus control. The VERO monolayers without any treatment served as cell controls. Calculation of the TCID50 was based on the Reed and Muench formula1
Plaque reduction assay
Dulbecco’s agar overlay plaque technique2 was modified to assay the effects of plant extracts on measles virus replication. The VERO cells were grown in plastic disposable culture bottles (25cm2 growth area) until confluent monolayers had formed. Growth medium was withdrawn from the cultures, and the monolayers washed twice with calcium and magnesium-free phosphate buffered saline solution. One ml of Eagles MEM with virus containing the equivalent of 70-80 PFU’S (calculated from the accompanying literature from the supplier) was added. The inoculated cultures were incubated at 370c for 2hrs. The cultures were overlaid with an agar preparation consisting of 2ml of 2% agar (Oxoid) at 460c and 2ml of solution A made in the following way: 100ml of 10 x Eagle’s MEM (with Earle’s BSS), 10ml of 3% Glutamine, 50ml fetal bovine serum (FBS), 25ml of 7% NaHC03, 2ml Pen/Strept/Amphotericin-B, 305ml sterile distilled water. This was the “first overlay”. Equal volumes of extract were incubated with virus for one hour and then inoculated, allowed to adsorb for one hour before being overlaid with the first overlay. The cultures were incubated at 370c for 5-7 days. To facilitate plaque counting, a “second overlay” was applied. This second overlay was of the same composition as the first except that it contained 1% Neutral red also. A plaque reader was used to view the plaques. Control flasks contained virus alone, extract alone. RESULTS i) Phytochemical studies Characterization of 27.135D as skimmianine Compound 27-135D recrystallized from methanol. The 1H-NMR (90MHZ, CDCl3, TMS internal standard) had features typical of skimmianine. The three singlets (3H, each at δ4.03, 4.12 and 4.4 are due to three methoxy groups. The two furano protons are indicated by two doublets at δ7.0 and 7.57 and the two doublets at 7.2 and 7.9 indicate the presence of two ortho aromatic protons at C6 and C5, respectively. The melting point, determined on a Koffler hot stage appraratus was within the range for skimmiamine (176-178oC). Comparison was made by superimposition of 1H3-NMR spectrum over authentic spectra of skimmianine .
ii) In vitro activity against measles virus Cytotoxicity assays
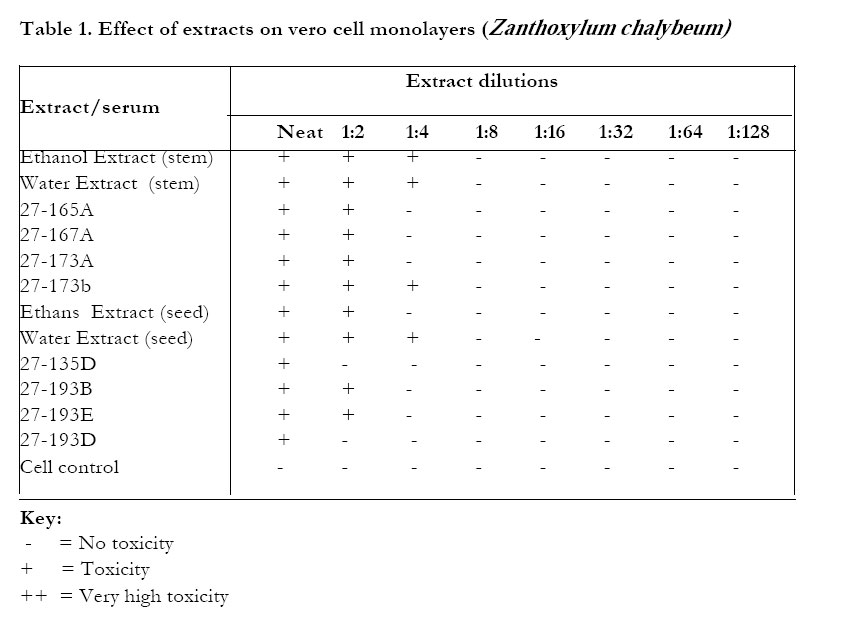
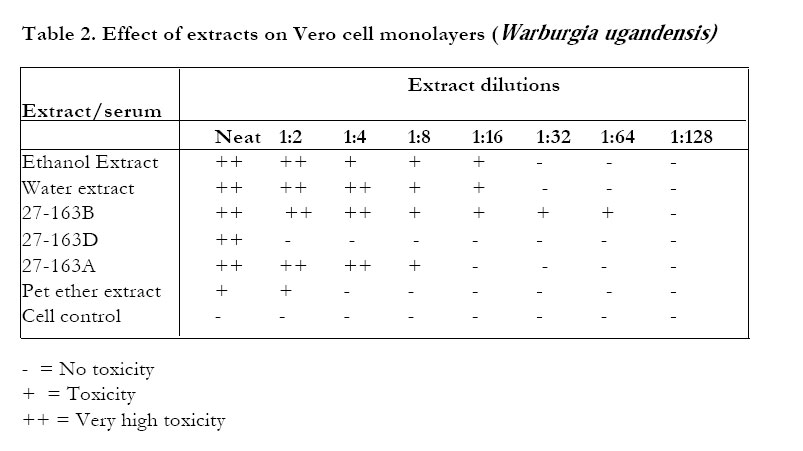
On VERO cells (Table 1) Zanthoxylum chalybeum extracts were essentially non-toxic - most extracts being non-toxic to the cells at the dilution of 1:8 from the stock concentration. Skimmianine (27-135D) was the least toxic of the fractions/components of Zanthoxylum chalybeum, being nontoxic even at the neat concentrations (Table 1). Warburgia ugandensis extracts were generally more toxic (Table 2). 27-163B was toxic even at a dilution of 1:64 from the stock concentration. The least toxic extract of W.ugandensis to the vero cells was 27-163D (the aqueous fraction of the ethanolic extract).
Antiviral studies
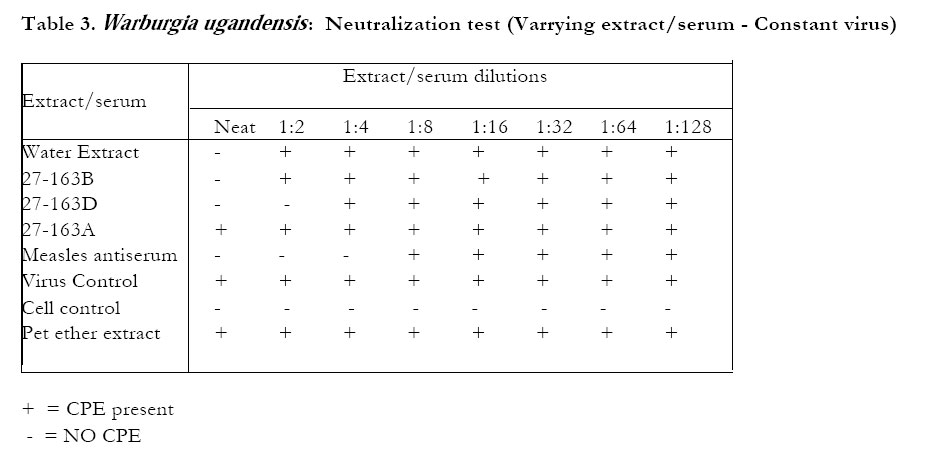
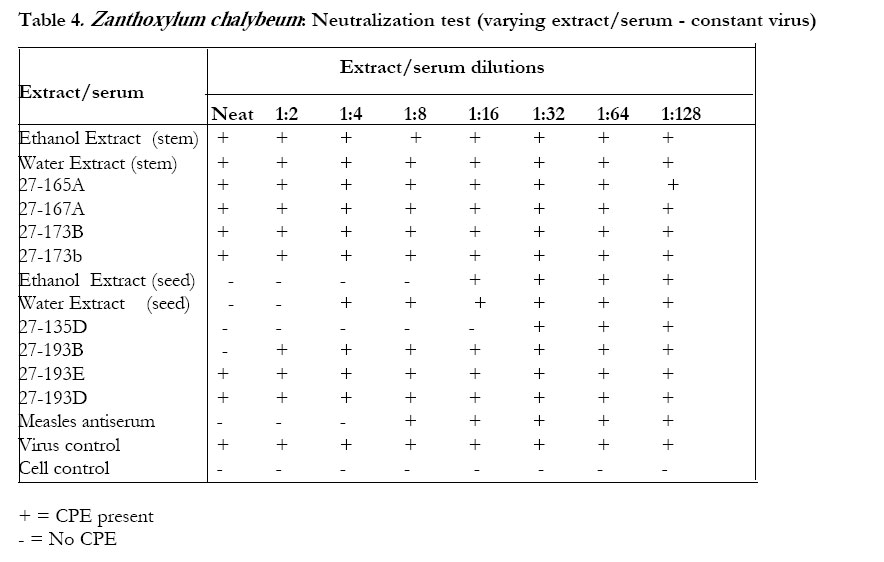
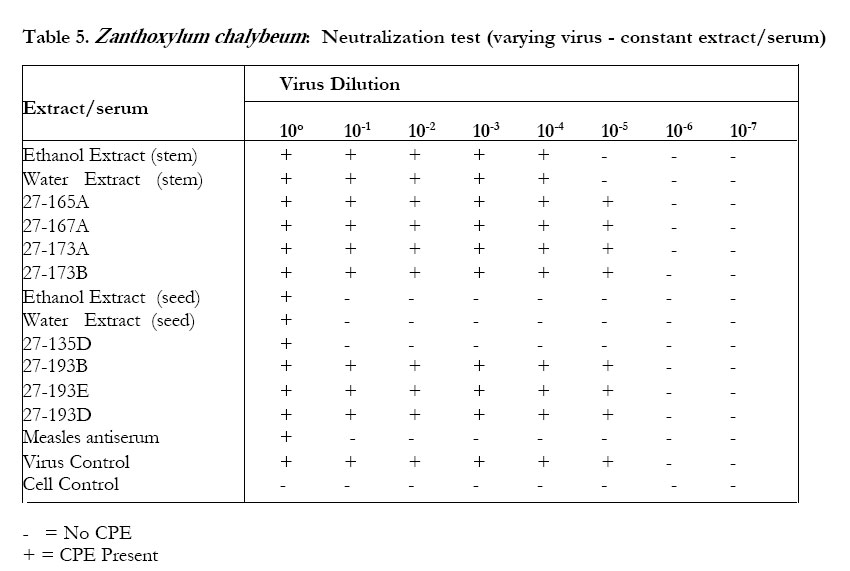
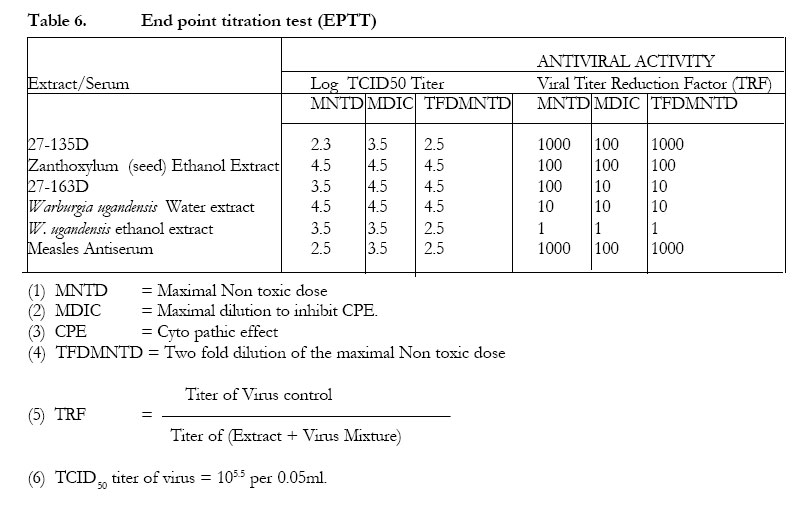
The petroleum ether extract ofWarburgia ugandensis, fraction 27-163B and the crude ethanolic extract did not inhibit CPE in the neutralization test (Table 3). The water extract and the aqueous fractions of the ethanolic extract (27-163D) showed some inhibitory effect on CPE formation, especially at the MNTD. The titer reduction factor (TRF) values were: water extract, 10; ethanolic extract,1; 27-163D, 100. All the stem bark extracts (alcohol, water and pet ether) of Zanthoxylum chalybeum did not inhibit CPE in the neutralization test. The alcoholic extracts of the seed and the alkaloid 27-135D (skimianine) inhibited CPE (Tables 4 and 5). The titer reduction factor (TRF) values were: ethanol extract (seed), 100; 27-135D, 1000 (Table 6). DISCUSSION This study did not demonstrate significant antiviral activity against measles in extracts derived from Warburgia ugandensis. Therefore, if W. ugandensis has any medicinal value at all against measles, it may be by some other mechanism other than direct antiviral activity (e.g. indirectly by conversion of inactive precursor to active form). This was rather strange, considering that the plant has been reported to contain several compounds with interesting biological activities including antifungal activity attributed to polygodial4,5,6 and trypanocidal activity attributed to the sequiterpene muzigadial7. Some sesqiterpenes from other plant sources have been reported to possess antiviral activity8. Previous studies have also reported cytotoxic properties, with a suggestion that some of its components could have anticancer properties . There have been no reports of antiviral activity of extracts of W. ugandensis. The seed extracts of Zanthoxylum chalybeum, however, exhibited some antiviral activity in vitro in this test system. The stem bark extracts did not. This result suggests that seed extracts are to be recommended rather than stem bark extracts for the treatment of measles. This may also be enviromentally advantageous because there is no harm to the trees (as compared to the effects of debarking when stem bark is used). Traditionally, water extracts of seeds (Z. chalybeum) and stem are prepared and taken orally. The antiviral activity of the seed extracts of Z. chalybeem seems to be attributable to the alkaloid skimmianine that is found in abundance here. The claimed benefit of Z. chalybeum extracts in treating measles may therefore be due to the antiviral action of skimmianine. There is need for further studies on this alkaloid because the demonstration of potency against the measles virus by extracts of one of the plants may indicate a possible use of the plant extracts for the treatment of other viral diseases such as human immunodeficiency virus (HIV), and other immunocompromised states (like in low-grade malignant non-Hodgkin lymphoma) where measles has been reported to be fatal9. It is not yet clear whether measles is the only virus that is susceptible to skimmianine since viruses respond differently to various plant extracts10,11,12. Even viruses in the same group react differently. For instance, in a study of poliomyelitis and cocksackie viruses13, found that the same product did not inhibit their multiplication equally. While it has often been recommended that a plaque reduction assay (PRT) is the best for assaying antiviral agents13, it was not possible to do the PRT in this study due to the difficulty of eliciting plaque formation in the measles virus in our test system. Previous studies in Africa14 and elsewhere12, 15, 16 have indicated that plants are potentially good sources of antiviral compounds. The demonstration of antiviral activity in Z. chalybeum seed, and in skimmianine in particular, in this study, could be a step forward, since our antiviral armamentarium is quite small and limited to the treatment of only a few specific viruses. This is especially due to the nature of the infectious viral agents, which totally depend upon the cell they infect for their multiplication and survival, so that many compounds that may cause the death of viruses are also very likely to injure the host cells that harbor them. It remains still to be established whether the in vitro findings of activity in this study can be of any significance in vivo. This is an area for further investigation.
ACKNOWLEDGEMENTS
This study was funded by the GTZ veterinary project based at the Faculty of Veterinary Medicine, Makerere University. The cell culture and antiviral studies were done at the Virus research Centre at KEMRI, Kenya. We are grateful to the director (VRC), Dr. Tukei, for permission to do the work in their laboratories. The phytochemical analysis was partly done at the Department of Chemistry, Addis Ababa University (AAU). We are grateful to Prof. E. Dagne and NAPRECA for part support to enable the D. Olila to do the study in Ethiopia. We thank Ms. Tenaye Asrat (AAU) and Mr. Muli (KEMRI) for excellent technical assistance.
REFERENCES
Copyright 2002 - Makerere Medical School, Uganda
The following images related to this document are available:Photo images[hs02029t5.jpg] [hs02029t6.jpg] [hs02029t4.jpg] [hs02029t1.jpg] [hs02029t2.jpg] [hs02029t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}