 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Health Sciences, Vol. 7, No. 3, June, 2007, pp. 166-175 Burkitt's lymphoma in Africa, a review of the epidemiology and etiology Jackson Orem a,b, Edward Katongole Mbidde a, Bo Lambert b, Silvia de Sanjose c & Elisabete Weiderpass b,d a Uganda Cancer Institute, Mulago Hospital and the Makerere University School of Medicine, Kampala, Uganda. Corresponding author: Jackson Orem, c/o Elisabete Weiderpass Department of Medical Epidemiology and Biostatistics Karolinska Institute P. O. Box 281, SE-171-77 e-mail: jacksonorem@ki.se or eliwei@ki.se Phone (EW-mobile) +35 8 40 845 3406 Code Number: hs07033 Abstract Burkitt’s lymphoma (BL) was first described in EasternAfrica, initially thought to be a sarcoma of the jaw. Shortly it became well known that this was a distinct form of Non Hodgkin’s lymphoma.The disease has given insight in all aspects of cancer research and care. Its peculiar epidemiology has led to the discovery of Epstein Barr virus (EBV) and its importance in the cause of several viral illnesses and malignancies.The highest incidence and mortality rates of BL are seen in Eastern Africa. BL affects mainly children, and boys are more susceptible than girls. Evidence for a causal relationship between EBV and BL in the endemic form is fairly strong. Frequency of association between EBV and BL varies between different patient groups and different parts of the world. EBV may play a role in the pathogenesis of BL by deregulation of the oncogene c-MYC by chromosomal translocation.Although several studies suggest an association between malaria and BL, there has never been a conclusive population study in support of a direct role of malaria in causation of BL.The emergence of HIV and a distinct subtype of BL in HIV infected have brought a new dimension to the disease particularly in areas where both HIV and BL are endemic. BL has been reported as a common neoplasmin HIV infected patients, but not in other forms of immuno-depression, and the occurrence of BL seems to be higher amongst HIV positive adults, while the evidence of an association amongst children is still disputed.The role of other possible risk factors such as low socio-economical status, exposure to a plant species common in Africa called Euphorbiaceae, exposure to pesticies and to other infections such as schistosomiasis and arbovirus (an RNA virus transmitted by insect vectors) remain to be elucidated. Introduction Burkitt lymphoma’s (BL) was described more than five decades ago by Dr Denis Burkitt in Uganda. However Sir Albert Cook, a missionary doctor in Uganda in 1887, had reported seeing children with similar features earlier. Despite all the initial work, BL still remains an elusive disease, which has provided many lessons on viral carcinogenesis and molecular oncology.1-3 Over recent years the incidence of BL has increased in the endemic areas in Africa, overlapping with the epidemic of HIV and malaria in the region. This review aims at providing the current status of understanding of epidemiology and etiology of Burkitt’s lymphoma. Burkitt lymphoma clinical variants: There are several forms of BL according to its geographic distribution, incidence magnitude and risk factors (Table 1). Endemic BL (eBL) is the disease originally described by Burkitt and largely found in Africa, characteristically affecting the facial skeleton in children between ages two to nine. Sporadic Burkitt’s lymphoma (sBL) is the form subsequently described outside the African region, but morphologically similar to eBL and affecting mainly abdominal viscera; it can be detected at any age and no specific co-factor has been described. A third subtype of BL has been proposed based on its association with HIV infection. Though well described in the developed world and known among HIV positive adults in Africa, the childhood form of the disease among HIV positive children has not been well characterized. HIV associated BL can be identified in any geographical area and at all ages and is of great importance especially in sub-Saharan Africa.4,5 Table 1. Overview of Burkitt's lymphoma clinical variants.
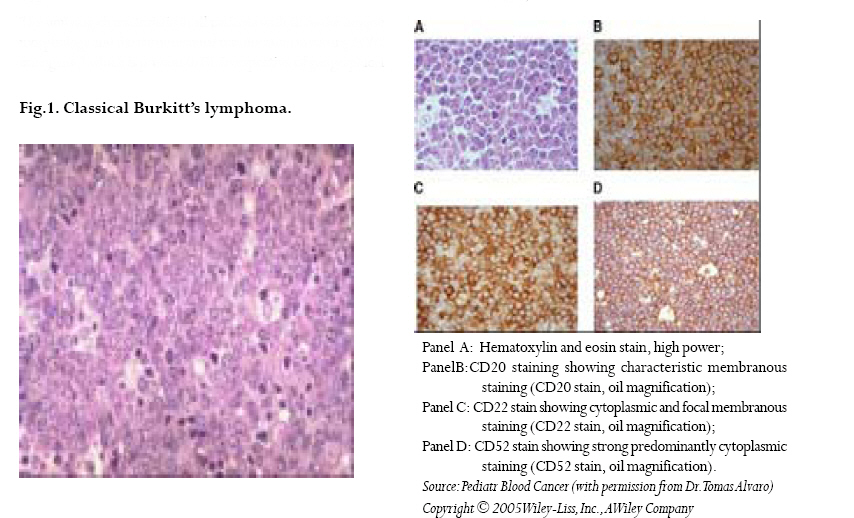
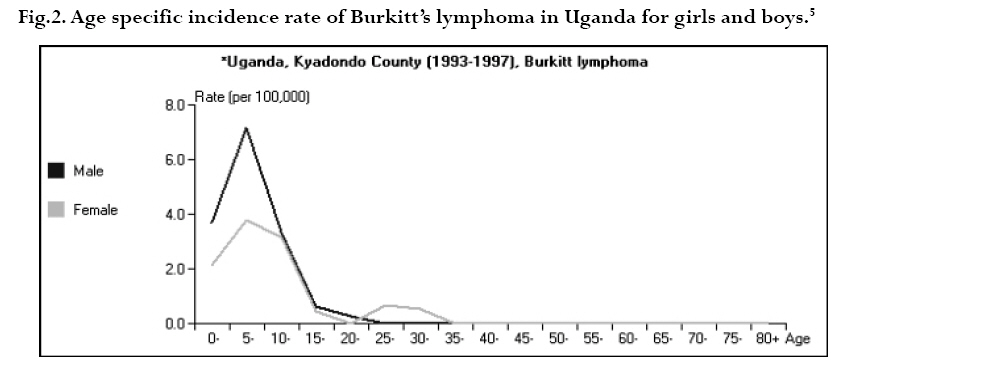
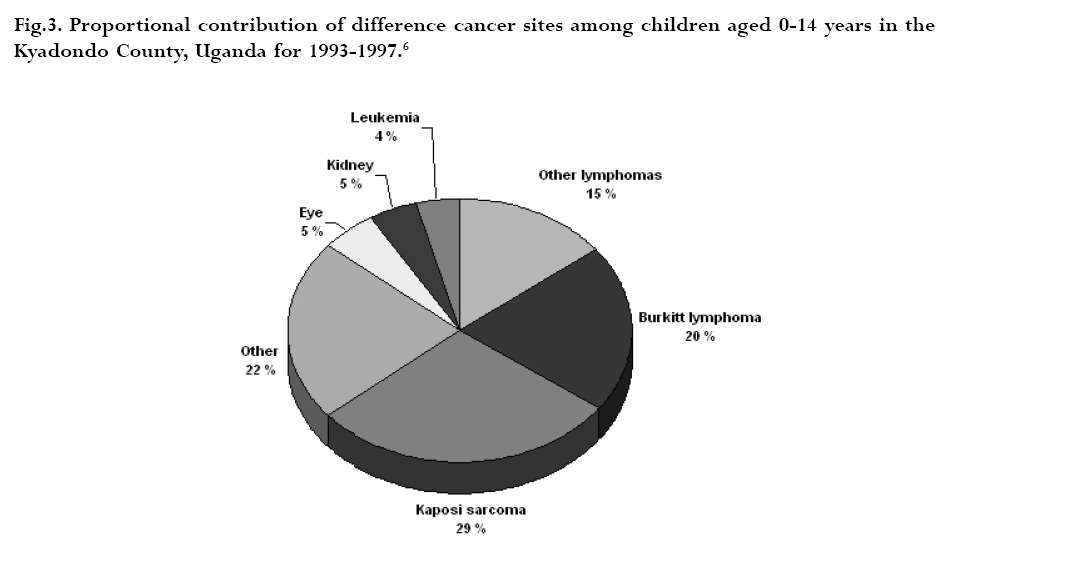
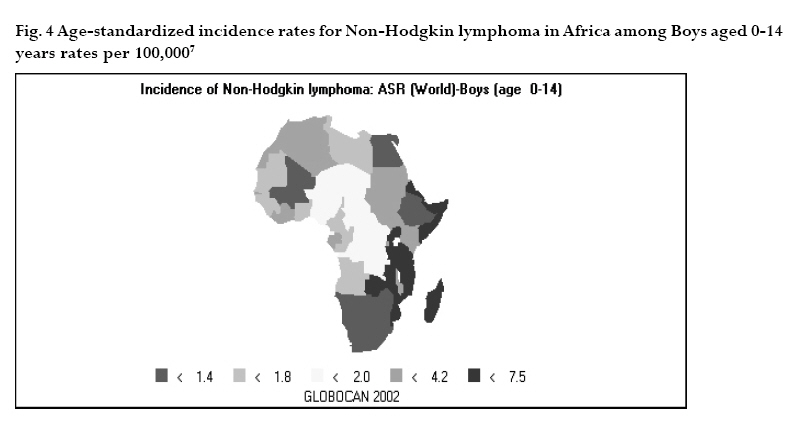
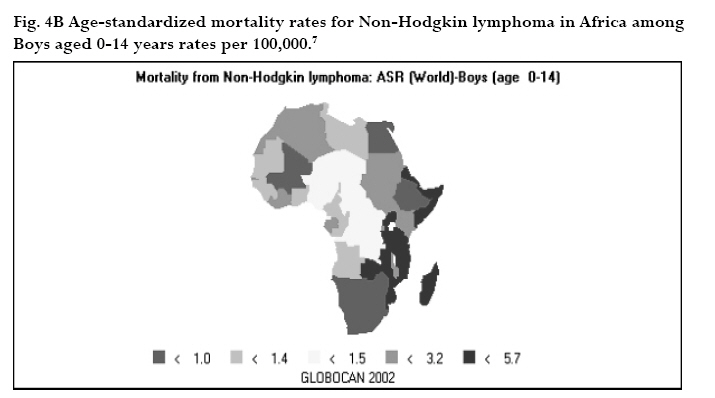
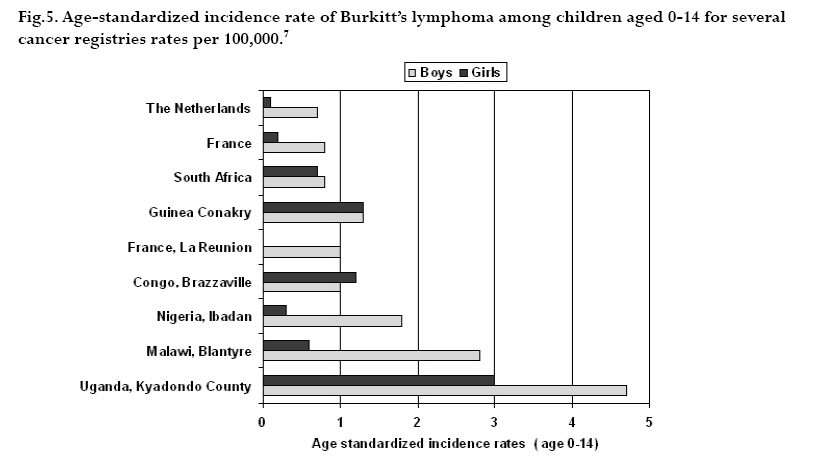
CNS = central nervous system Ig = immunoglobulin EBV = Epstein-Barr virus Sources: Cancer in Africa, 2003 5 & Cancer Incidence, Mortality and Prevalence Worldwide, 2004.6 BL is included in the World Health Organization (WHO) classification of tumours of haematopoietic and lymphoid tissues, with an ICD-O code of 9687/3 for the Burkitt’s lymphoma nodal modality and the ICD-O code 9826/3 for the Burkitt’s leukemia modality.4 Morphologically the WHO classification distinguishes between classical BL, generally observed in eBL and sBL; BL with plasmacytoid differentiation seen more commonly among immune deficient children and finally the atypical Burkitt or Burkitt-like variant. The unifying characteristic in all patients with BL is the unique morphology and the chromosomal translocation involving MYC oncogene,4 which is present in BL irrespective of geographical location, and immunodeficiency status. Burkitt’s cells are monomorphic cells with round nuclei, multiple nucleoli, and basophilic cytoplasm with a high rate of proliferation and spontaneous cell death. Cytoplasm lipid vacuoles are usually evident on imprints or smears.The rate of cell division is the highest of any known tumor noted by the very high mitotic figures. A starry-sky pattern is usually present, imparted by numerous benign macrophages that have ingested apoptotic tumor cells (Fig. 1). 2.The burden of Burkitt lymphoma in Africa BL which is endemic in Africa affects mainly children, and boys seem to be more susceptible than girls (Fig. 2).5 Data form Globocan 2002 has been used to estimate the proportional contribution of different cancer sites among children aged 0-14 years in the Kyadondo County, Uganda for 1993-1997 (Fig. 3).6 The most common malignancy was Kaposi sarcoma accounting for 29% of all cancers, followed by BL accounting for 20% of all pediatric cancers and in third position all other lymphomas accounting for 15% of all cancers. The geographical distribution of BL can be derived from the general pattern of incidence and mortality of Non-Hodgkin Lymphoma (NHL), as BL is the single most common entity contributing to NHL in Africa. Figure 4a, 4b shows the age-standardized incidence and mortality rates for NHL in Africa among boys aged 0-14 years. The highest NHL incidence and mortality rates are seen in Eastern Africa with estimated incidence rates over 7.5 per 100.000 and mortality rates over 5.7 per 100.000. Incidence and mortality show exactly the same regional pattern. A similar pattern is observed for girls.6 Ad hoc studies indicate that BL is more common in areas where malaria is endemic. This geographical variation was investigated by Burkitt, who pointed out the association between humidity, latitude and heat as factors linked to the endemicity of malaria and of BL (see below). However, when looking at the incidence data by country, it can be seen that BL is more common in Eastern Africa as compared to other African countries where malaria is endemic (Fig. 5). Within the Cancer Registries available in Africa, the incidence of BL is highest in the cancer registry from the Kyadondo county, Uganda, with an age-standardized rate (ASR) per 100.000 of 4.7 for boys and 3.0 for girls (in all 90 cases reported in a five-year period of registration).The second ranking registry is Malawi with ASR per 100.000 of 2.8 for boys and 0.6 for girls. Incidence rates of BL in Mali, Nigeria, Congo and The Gambia are lower than those reported in Uganda but substantially higher than those observed in other African Regions (Table 2) or in Europe (Fig. 5). African countries in the lower range of incidence show values around 1 case per million which are lower than those observed in France (8 cases per million) or in The Netherlands (7 cases per million). Country differences in reporting practices or real country-based differences are of relevance in order to evaluate the role of co-factors inducing BL.5 In the recent decades, there has been an increase in incidence rates of many malignancies, some of which can be related to changes in lifestyle and the HIV epidemic. Among the HIV related malignancies, NHL and BL have been most affected. In Uganda, BL incidence rates have was 6.6 years.7 Although part of the increase could be been reported to increase from 9.5 per 106 in 1960-71, explained by better case ascertainment, HIV is likely to to 34.3 per 106 for the period 1991-97. The sex ratio have largely contributed to this increase. male: female were 1.6 for the first period and 1.5 for the later period.The mean age at diagnosis for both periods
Table 2.Age standardized incidence rates of Burkitt’s lymphoma in Africa by gender rates per 100,000.
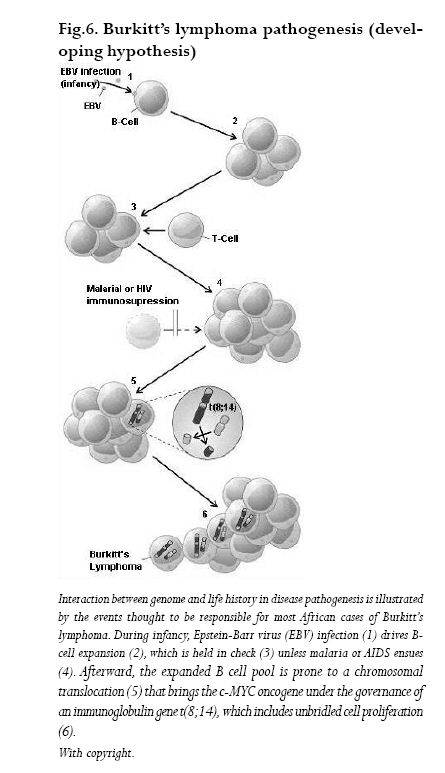
ASIR= Age standardized incidence rates Sources: Cancer in Africa, 2003 5 & Cancer Incidence, Mortality and Prevalence Worldwide, 2004.6 Molecular signatures of BL BLs cells derive from lymph node germinal center regions where rapidly proliferating B-cell blasts (centroblasts) differentiate into centrocytes following hypermutation of the immunoglobulin gene variable (IgV) region and transform into malignant BL and/or DLBCL (Diffuse large B cell lymphoma). Several recent reviews have discussed the molecular biology of BL,8,9 and the central role of c-MYC deregulation in the pathogenesis of the disease.10 Chromosomal rearrangement of the proto-oncogene c-MYC is the genetic hallmark of BL: over 80% of BL cases have a translocation of MYC at band q24 from chromosome 8 to the Ig heavy chain regions on chromosome 14, t(8;14). Less frequently rearrangements translocate c-MYC to a position close to the antibody genes in chromosome 2 or 22. In every case, c-MYC finds itself in a region of vigorous gene transcription (Fig. 6).These translocations contrib-ute to lymphoma genesis through alterations in the cell cycle regulation, cellular differentiation, apoptosis, cellular adhesion, and metabolism. In endemic BL cases the c-MYC translocation involves the heavy chain joining regions while in sBL or in HIV-associated-BL the translocation involves the Ig switch region. MYC translocations are not exclusive of BL and can also be detected in other lymphoma subtypes such as T-cell lymphomas or multiple myeloma. The WHO classification requires for a diagnosis of BL a high growth factor, with Ki67 staining over 99% and evidence of c-MYC rearrangement when the analysis is possible. Other genes may be responsible for the evolution and progression of BL such as p53 mutations, methylation of death-associated protein kinase (DAP-kinase), or changes in other cellular proteins such as p16INK4a or p15INK4b through gene hypermethylation.11 The differences between high and low incidence regions and HIV associated BL could be partly explained by different molecular changes in the BL subtypes. The understanding of the genetic markers of the disease may elucidate etiological mechanisms inducing BL. Risk factors for Burkitt’s lymphoma EBV Discovery of EBV was a direct outcome of the clinical description of BL.1,2 The restricted geographic distribution of BL in Africa led to a hunt for a possible viral cause and eventually the discovery of a novel human herpesvirus.12-15 EBV is a lymphotrophic gamma human herpesvirus widely spread among humans, and was the first virus to be associated with a human tumor. Transmission of EBV takes place early in life and is associated with human contact through saliva.16,17 In Uganda, children with higher baseline titers to EBV antigens are at a higher risk of developing BL, and higher antibody levels were detected many years before BL diagnosis.18 The capacity of EBV to stimulate B-cell proliferation inducing malignant phenotype strongly supports an etiological role of EBV in BL.19 In 1997, the International Agency for Research on Cancer concluded that there was sufficient evidence for the carcinogenicity of EBV in the causation of BL.20 EBV may play a role in the pathogenesis of BL by deregulation of c-MYC though (a) potentiation of c-MYC activity and clonal expansion, (b) direct mutagenesis and (c) immune inactivation leading to escape of malignant clones. EBV is known to transform resting B cells into latently infected lymphoblastoid cells. Constitutive expression of the latency state is evidenced by presence of latent proteins consisting of six EBV nuclear antigens (EBNAs 1,2,3A, 3B, 3C and –LP) and three latent membrane proteins (LMPs1, 2A and 2B).The expression of non-coding poly adenylated small RNAs, EBER1 and EBER2, called latency III, are observed.The expansion of the B-cell pool as a result of the co-infection with EBV is regulated by the immune system; immunosupression caused by infections such as HIV and malaria could lead to chronic stimulation of B-cells increasing the risk of c-MYC translocation and other mutations. Figure 6 illustrates the relationship between EBV, malaria infection and the genetic changes that drive the B-cell into BL. Malaria Infections with P. falciparum in holoendemic equatorial Africa may occur throughout the year, the first infection usually occurring during the first year of life, peaking at around age 5 and leveling off after age 10. After repeated infections tolerance to P. falciparum infection may appear after age 8 years. Although P. falciparum is not considered as an oncogenic agent, its role in the development of BL has been suggested by the shared geographic distribution of eBL and holoendemic malaria. Both diseases show highest incidence rates in the lymphoma belt or malaria belt, an area around the equatorial Africa (+/-10º) where average temperature exceeds 15.5ºC and have yearly rainfall higher than 50 ml. Endemic BL outside Africa is seen in other malaria holoendemic areas such as Papua New Guinea while it is also observed in Brazil where malaria is not holoendemic.21 The fact that eBL is associated to EBV infection has induced research towards a joint mechanism of action between P. falciparum and EBV infection. The review by Rochford et al.22 summarizes the pathways by which both infections could induce eBL: malaria could disrupt immune response against EBV, leading to BL. Further, reduction of BL incidence was observed in areas where malaria was eradicated, and BL onset delayed among subjects migrating from a low to high-risk malaria areas. Co-variation between susceptibility to malaria and BL has been documented. Some haemoglobinopathies, such as sickle cell trait or haemoglobin trait, have a protective effect against malaria and BL.23 Children living in malaria areas have high immunoglobulin levels, a marker of an increased immune reactivity to P. falciparum. However, there has never been a conclusive population study in support of a direct role of malaria in causation of BL. In Tanzania, where BL rates are high,24 intervention with Cloroquine chemoprophylaxis against P. falciparum infection in children diminished by three fold the incidence of BL, but this was followed shortly thereafter by a rebound in the BL incidence rates despite chemoprevention.Thus, there is still need for a population model to understand the specific role of P. falciparum in the causation of BL: the effect of age at first P. falciparum infection, percent of cells infected and recurrence rates of malaria are yet to be understood. Arbovirus, a RNA virus transmitted by insect vectors, may play a role as final cofactor in the etiology of eBL by inducing cell growth and B-cell hypermutation,23 but data-supporting hypothesis are scarce. HIV BL has been reported as an common neoplasm in HIV infected patients, but not in other forms of immuno-depression. HIV associated BL display an activation of c-MYC by chromosome translocations that show structural similarities to those found in patients with sporadic BL. Nonetheless, most AIDS related BLs in Western countries are EBV negative,whereas in Africa they are strongly associated with EBV. However, the association BL-HIV has been detected at a lower rates in Africa as compared to that observed in the western world.25,26 Recent data show a gradual rise of BL incidence rates as well as other HIV related tumors. The impact of AIDS on the incidence of Non-Hodgkin’s lymphoma in Africa is just beginning to be systematically studied and changes in the profiles in Sub Saharan Africa are emerging.27,28 Still, BL in children are more likely to be of the endemic form than the one associated to HIV, as shown by two studies from Uganda,29,30 although another study31 have identified a strong association between HIV and BL in children in Kampala.The rise in childhood lymphoma and especially of BL in some regions of Africa is not clearly understood but suggests a possible association with HIV. Poor survival of children infected with HIV perinatally (only 34% survive to the age of 3 years)32 has been suggested as one of the explanations for the lack of association found in some studies. In adults, 3-fold increase in BL (66% HIV+) between 1992-1996 was reported in a survey in Kenya,33 a finding further supported by data from Uganda indicating an increase in adult BL as a subset of NHL in HIV infected adults.29,34 Other factors Herbal exposure:The sap of the milk bush (Euphorbia tirucalli spurge) and other Euphorbiaceae species are possible environmental risk factors for BL due to their ability to activate the viral replication cycle in the latent phase of EBV-infected cells. The milk bush is a succulent plant containing rubbery, white latex, which grows as shrub or small trees in many parts of tropical Africa (Fig. 7). Extract of Chinese and African Euphorbiacae plants, including several Croton species were able to enhance the EBV-induced transformation of human lymphocytes.40 This effect of the plant sap was attributed to its content of croton oil, the parent compound of the tumour promoter TPA (12-Otetradecanoylphorbol-13-acetate) which enhances EBV-induced transformation of human lymphocytes.41 TPA induces the expression of the viral immediate-early protein BZLF1 through a posttranslational mechanism involving protein kinase C (PKC) - dependent phosphorylation.42 Low concentrations of crude E.tirucalli latex extracts is capable of reactivating the EBV lytic cycle in a dose-dependent manner through activation of a PKC pathway.43 E. tirucalli contains diterpene esters including 4-deoxyphorbol ester.44,45 In a study of human cord-blood B-lymphocytes exposed concomitantly to EBV and 4-deoxyphorbol ester purified from E. tirucalli c-MYC expression and chromosome aberrations typical for BL were induced, such as translocations involving the long arm of chromosome 8. These changes were not induced by EBV or 4deoxyphorbol ester alone.46 These reports suggest a possible role of extracts E. tirucalli and other Euphorbiaceae species in the development of BL through activation of the lytic cycle of EBV-infected cells, and promoting a sustained production of infectious virus. This role of Euphorbiaceae species as possible environmental co-factors in the pathogenesis of eBL is supported by the finding from Malawi 47 where the plant is found more often at the homes of eBL patients than in those of controls. Euphorbiacae plants are used traditionally in many cultural activities in tropical Africa, including the Lake Victoria Basin, such as religious, wedding and birth of twin ceremonies, human (to treat sore throat, headache, diarrhoea and wounds) and veterinary medicine, as a poison for fishing, and for construction of shrines and protective fences,44,45,48-50 and may be in children’s plays.43 Other environmental factors to be considered in Africa should include schistosomiasis21 and exposure to pesticides: for both, the evidence supporting this association is sparse. Summary There is evidence for causal relation of EBV infection and BL at a population level in equatorial Africa, although the mechanisms of carcinogenesis are not completely understood. The role of malaria and HIV in the development of BL is not clearly elucidated.The role of socioeconomic factors, infectious agents leading to disturbance of immunity such as schistosomiasis, viral infections such as arboviruses, and exposure to environmental factors, such as exposure to E. tirucalli and other Euphorbiaceae, need to be further elucidated. Acknowledgements This work was supported by SIDA/SAREC (Sweden) though a grant for the collaboration between the Karolinska Institutet, Stockholm, Sweden and Makerere University, Kampala, Uganda, ICRETT fellowship number ICR R 06/139, and the Cancer Registry of Norway, Oslo, Norway. Prof. Bo Lambert was supported by an Expert Transfer Fellowship from the International Agency for Research on Cancer, Lyon, France. Dr. Silvia de Sanjose was supported by CIBERESP (Spanish Ministry of Health). All authors declare no conflict of interests. References:
Copyright © 2007 - Makerere Medical School, Uganda The following images related to this document are available:Photo images[hs07033f6.jpg] [hs07033f4a.jpg] [hs07033f5.jpg] [hs07033f3.jpg] [hs07033f7.jpg] [hs07033t1.jpg] [hs07033t2.jpg] [hs07033f1.jpg] [hs07033f2.jpg] [hs07033f4b.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}