 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Health Sciences, Vol. 7, No. 4, December, 2007, pp. 190-196 A study of physico-chemical interactions between Haemophilus influenzae type b and meningococcus group C conjugate vaccines. 1 Robert B. D. Otto*, Dennis T. Crane and Barbara Bolgiano 1Division of Bacteriology, National Institute for Biological Standards and Control, Blanche Lane, South Mimms, Potters Bar,
Hertfordshire EN6 3QG, United Kingdom. Code Number: hs07038 AbstractBackground: Haemophilus
influenzae type b (Hib) and Meningococcal group C (MenC) conjugate vaccines, which protect against
otitis media, bacteremia and invasive diseases, including pneumonia and meningitis, are attractive candidates for combination, since they
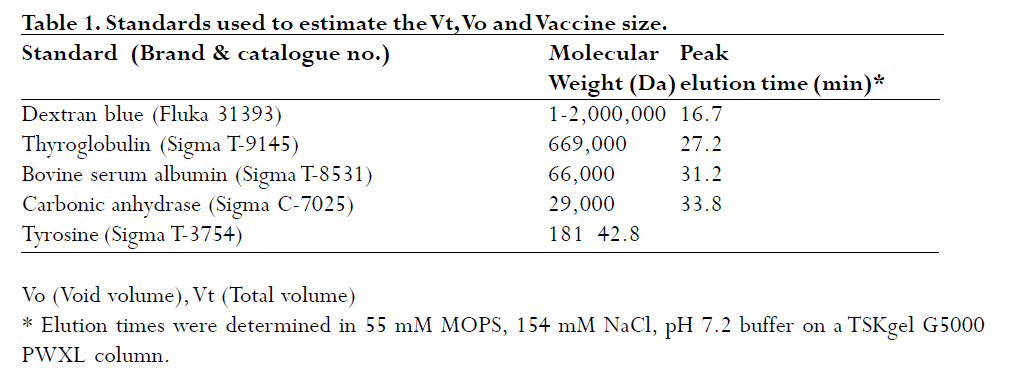
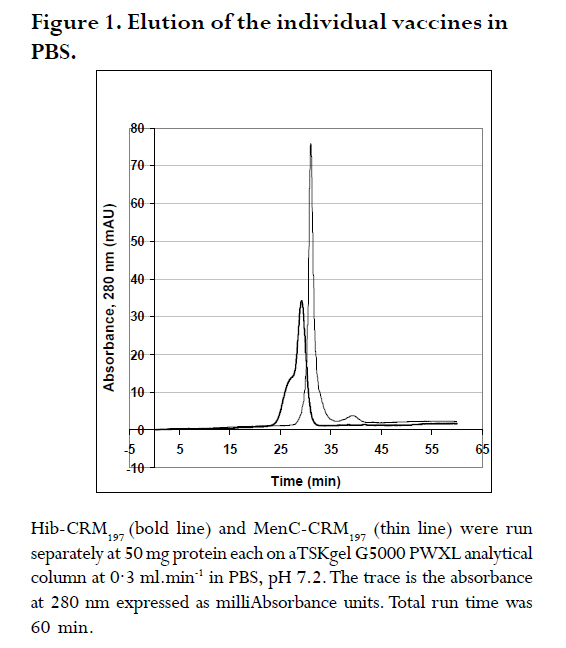
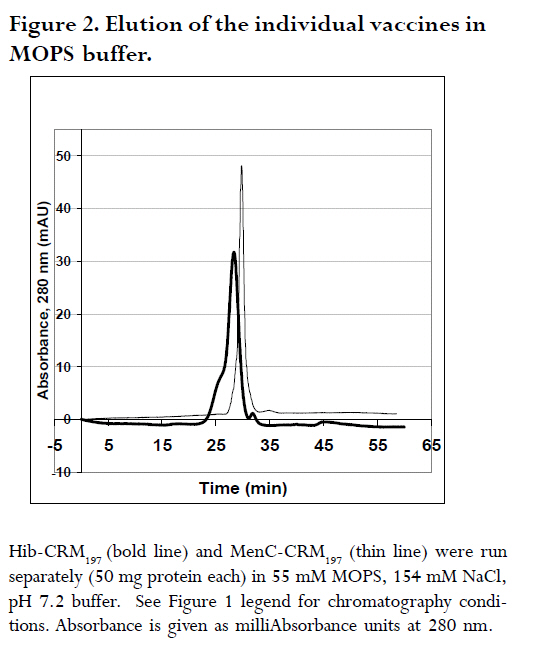
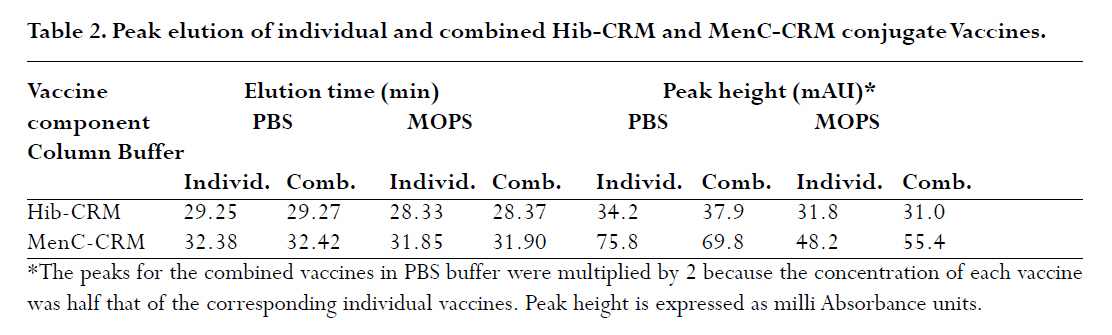
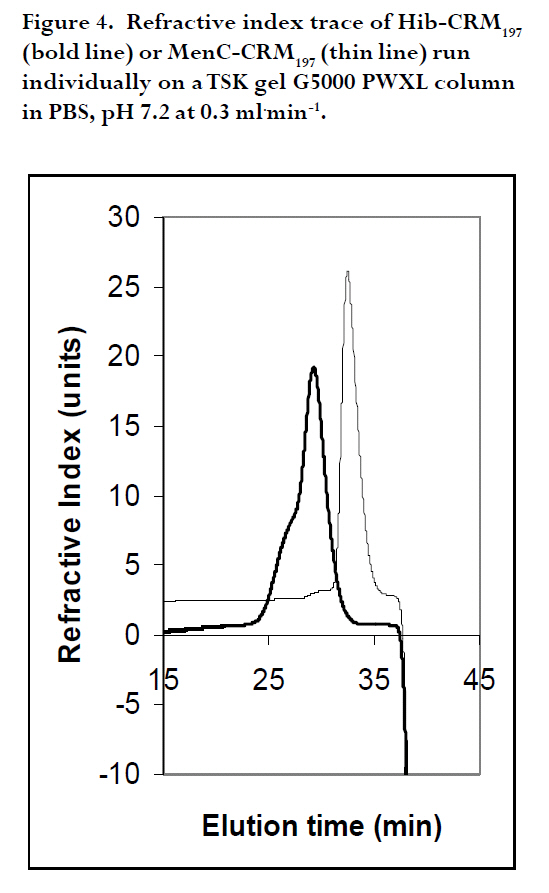
are both administered to infants and children. A Hib-MenC combination booster at 12 mo has recently been introduced in the U.K. Introduction The effectiveness of vaccines to eliminate Haemophilus influenzae type b (Hib) disease has been well documented. Recent reports of overwhelming success in Kenya and The Gambia, two African countries implementing childhood Hib saccharide-protein conjugate vaccine immunization, have shown disease incidence decreasing from 60-200 cases /100,000 children prior to vaccination to 0-26 cases /100,000 children following vaccination1, 2. Control of N. meningitidis-derived invasive infections in the Meningitis Belt of Africa has primarily been limited to the use of antibiotics and polysaccharide vaccine, when available, with case incidences rising to 500/100,000 in epidemic years3; in 2006-2007, a significant increase over baseline (non-epidemic) levels has been observed with more than 48,000 cases and 3,400 deaths reported through enhanced surveillance programs, leading the Meningitis Vaccine Project to speculate that regions may be nearing an epidemic cycle4. In the United Kingdom, where this study was performed, as a result of successful immunization programs for Hib (commenced in 1992 and group C meningococcal (MenC) conjugate vaccines (commenced in 1999), the incidences of both diseases in England and Wales have dramatically decreased from 22.9 to 0.65 cases Hib/100,000 children in 1998, 6 years following the start of Hib vaccination, and, from 4.1 to 1.4 cases MenC disease/100,000 vaccinated5, 6, although it has been recognized that administration of a booster dose of vaccine to toddlers would increase vaccine protectiveness in this and other age groups even further7, 8. Based on the proposal of the U.K. Joint Committee on Vaccination and Immunisation9,, the Department of Health introduced a Hib/MenC combination booster dose to toddlers in 200610. Hib and Men C conjugate vaccines are attractive candidates for combination, since they are both administered in the primary infant vaccination schedule in the U.K11. However, particular vaccine combinations containing acellular pertussis (aP) components have been associated with deleterious effects. The widespread use of a vaccine DTaP-Hib combination vaccine has resulted in a lowered immunogenicity of the Hib component5, 12, 13 and increases in Hib disease in multiple age groups 7. WHO Guidelines advise on the non-clinical evaluation of potential physical interactions between components 14-16, as well as clinical studies to verify the safety and immunogenicity. The possibility exists for the Hib and Men C components in the combination to interact with other antigens, formulation salts, excipients or adjuvants, rendering one or both of the antigens ineffective. Plausible mechanisms specific to the conjugates may involve the loss of stability or solubility due to association or aggregation of vaccine saccharide or protein components 17, 18 depolymerisation or hydrolysis of saccharide bonds 19, 20 resulting in shorter, insufficient epitopes and decrease in effective vaccine dose, or alteration in the adsorption and activity of adjuvants. These mechanisms may be affected by the formulation or buffer in which the vaccines are held18, 20, 21. This study assessed the viability of combining the two vaccines together, by characterizing Hib-CRM197 and MenC-CRM197 components through size-exclusion chromatography. In addition to assessing their integrity, the size of Hib and MenC conjugate components were assessed before and after combination to rule out antigen-antigen association, when in different buffers. Methods Vaccines: Both the Hib-CRM197 and MenC-CRM197 samples used in this study were the kind gift of their manufacturer. They consist of short (~15-20 repeating units) of Hib or MenC capsular oligosaccharides covalently conjugated to the protein carrier, CRM197, Cross- Reacting Material 197, which is a single amino acid non-toxic variant of the parent protein, diphtheria toxin22. The oligosaccharides, for both vaccines, are linear anionic chains consisting of either the Hib PRP disaccharide repeating unit (polyribosyl ribitol phosphate)23 or linked N-acetylneuraminic acid homopolymer with O-acetyl (OAc) groups at C-7 or C-8 residues24. CRM197 lysyl residues are conjugated either directly to Hib PRP or via a spacer to MenC oligosaccharide. The bulk conjugates were stored frozen: at -20°C for the Hib or -70°C for the MenC, which was used within 2 yr of manufacturing. Sample Preparation: Several mL of the bulk vaccines (Hib-CRM197 and MenC-CRM197) were dialysed in three changes of 1 L of normal saline (154 mM NaCl) at 4oC using 10,000 Da MW cutoff dialysis membrane (SpectraPor). Each first and last round of the dialysis lasted for 4 hr, whilst the 2nd round was performed overnight ≈14hr. Perculation was achieved by magnetically stirring the saline. Following dialysis, the concentrations of the protein in the vaccines in 1 mL solutions were measured using a Perkins Elmer Lambda 800 spectrophotometer (Perkin Elmer Inc., Waltham, MA, USA) using an absorption coefficient A279 nm (1mg/ml) of 0.757 cm×mg×mL-1 25 by measuring A278.6 nm - A450.0 nm. Samples were diluted to 500 mg protein×mL-1 with saline (154 mM NaCl) and kept at 4°C for the 1 mo duration of the study. High Performance Liquid Chromatography (HPLC/SEC): A Dionex DX600 HPLC system (Dionex, Sunnyvale, CA, USA) as used to perform size-exclusion chromatography. It was equipped with an eluent module, gradient pump, thermostatted column compartment set at 30 oC, UV/VIS-Detector (UVD 170U) with wavelengths set at 280 nm and 214 nm, and a SpectraSYSTEM RI-150 refractive index (RI) detector (Thermo Separation Products, Inc, Thermo Fisher Scientific, Inc., Waltham, MA, USA). The column used for separation was a TSKgel G5000 PWXL analytical (7.8 mm x 30 cm, ~12 mL volume) and PWXL guard column (Toso Haas, Tokyo, Japan). The flow rate was 0.3 ml×min-1 and running time was 60 min. Fifty μg protein (100 μl) of dialysed bulk Hib-CRM197 or MenC-CRM197 conjugate vaccines (each at a concentration of 500 ìg protein×mL-1) were injected and eluted in either a phosphate-buffered saline (PBS) buffer consisting of 50 mM sodium phosphate, 154 mM NaCl, pH 7.2 or 55 mM 3-(N-Morpholino)-propanesulfonic acid, 154 mM NaCl, pH 7.2 buffer (MOPS, Sigma) through the HPLC/SEC column. For the combined vaccines, equal volumes of the vaccines were mixed and loaded. The column eluent was monitored directly in a flow-cell for A214 nm and A280 nm followed by RI. The software employed was Chromeleon® (Dionex, Sunnyvale, CA, USA), for programming and running the chromatography and for analysis of the generated data. Data output is shown in terms of milliAbsorbance units (mAU) versus time. Duplicate experiments were performed, and results are expressed as the mean of two runs performed on different days. The standard deviation (n-1) of peak elution times of combined vaccines was determined to be ± 0.066 min (n=3) for samples run on different days. Vaccine size estimation and Column volume (Vt) and Void volume (Vo) Calibration: In order to obtain size estimations of vaccine components, and to determine the elution of void volume (Vo) and total volume (Vt) markers, standards of known molecular weights were chromatographed (Table 1) 26, 27. It was vital to get an idea about the size of the vaccine molecules being eluted so as to compare them with available data and verify whether the individual vaccines oligomerise/aggregate within themselves (intra-molecular associations), as well as to evaluate whether there was any inter-molecular association between the two components in the combination. These standard markers were eluted by the same buffers and their peaks of elution were also detected in the same way as for the vaccines in the chromatography. A Microsoft Office Excel plot of the natural log (ln) of the molecular weights of the standards versus peak elution obtained in MOPS buffer was used to estimate the molecular size of the main components of the vaccines. Results Size characterization of vaccine components HPLC/SEC-UV chromatographs from individual vaccines were superimposed for comparison of the peaks and are shown in the Figures 1 and 2. Distinct peaks for Hib-CRM197 and MenC-CRM197 were observed in phosphate and non-phosphate-saline buffers, pH 7.2. Hib-CRM197 eluted significantly earlier than MenC-CRM197, with a slight broadening of the peak and leading shoulder, suggestive of a higher degree of polydispersity than with the MenC-CRM197. The difference in the elution of the leading shoulder in PBS and MOPS suggests a slight aggregation or non-covalent oligomerisation of the Hib-CRM197 vaccine molecules. In contrast, the MenC-CRM197 eluted as a sharp peak with insignificant higher molecular weight elution. Interaction of individual vaccines in a combination The distinct elution profiles for the two vaccines allowed evaluation of the combination feasible by UV flow-cell monitoring only. Similar peak elution patterns before and after combination were observed for Hib-CRM197 and MenC-CRM197 (Figure 3 and Table 2) with only 0.02-0.05 min difference in elution time, within the determined experimental variation. This suggested there is minimal, if any, antigen-antigen association in the combined vaccines, although the saline buffers and pressures from chromatography could have disrupted weak interactions. Non-specific aggregation or association between Hib and MenC conjugate vaccines when combined together would have given newer earlier peak(s) apart from those of the individual vaccines and this would have been accompanied by a drop in peak heights of the individual vaccines and band broadening (poorer resolution) between peaks, as observed for many hetero- and homo- protein oligomers. In both PBS and MOPS, there was no significant difference between peak elution times of individual compared with combined vaccines. The vaccines (individual and in combination), elution times were different in the two different buffers, however, with the conjugates eluting earlier (0.5-0.95 min) in MOPS than in PBS buffer (See Figure 3 and Table 2), in line with the slight earlier elution of the markers (0.2 -0.4 min) in MOPS than in PBS. Evaluation of vaccine integrity To ensure that conjugate vaccines remain stable, limits are placed on the amount of free protein and free saccharide. Assays of total saccharide content and % free saccharide obtained in filtration, centrifugation, detergent-extraction, or bead separation supernatants are measured in bulk conjugates or final fills14, 28. It is possible that quantitative chromatography-based methods can be validated against standard colorimetric assays for measuring vaccine integrity, saving time and resources. The combined use of sensitive, calibrated RI (to detect protein+saccharide) and A280 nm (protein absorbance only) detectors makes it theoretically possible to resolve protein separately from saccharide; free protein and saccharide are also resolvable in the chromatographic system employed in this study. Free CRM197 carrier protein would be expected to elute after the main MenC-Hib peaks, and free Hib and MenC oligosaccharides and fragments would be observable in an RI signal prior to the Vt at 43 min. Neither the UV nor RI traces (Figure 4) of the individual Hib- and MenC-CRM197 indicates significant free protein or saccharide. The column void elution time (determined by blue dextran) was at 16.7 min and the total elution time (tyrosine) was at 43 min. Discussion The National Institute of Biological Standard & Control Laboratory (NIBSC), in its role as a national control laboratory, performs pre-clinical and batch release of Hib, MenC and other conjugate vaccines and biological therapeutics29. In addition to safety testing of toxoid and final product and identity assays of bulk saccharide and carrier protein components, bulk conjugates or final fills of conjugate and polysaccharide vaccines are evaluated by molecular sizing and saccharide-specific chromatography methods30. Molecular sizing by aqueous HPLC methods have proven to be sensitive to subtle changes in manufacturing and is an excellent indicator of consistency of production. In this study, the size and integrity of the different conjugates made by the same manufacturer were compared by molecular sizing. MenC-CRM197 eluted as a monodisperse monomer in PBS, pH 7.4 on a TSK 5000 polymer column, corroborating previous characterization of the conjugate, which has a weight-average molecular weight of 75 kDa based on SEC with light scattering detection 25. The Hib-CRM197 conjugate ran as a more polydisperse and larger-sized conjugate. Both conjugates had a high degree of integrity. Bond breakage and degradation of the individual vaccines into smaller particles would result in later peaks, peak broadening and appearance of trailing shoulders and/or drop in peak height/area of the individual vaccines, as seen in accelerated stability studies with conjugate 17, 18, 31, 32 or real-time studies with polysaccharide 33 vaccines. Hib-CRM197 and MenC-CRM197 do not appear to physically interact in solution, as measured by elution time and peak height, and their combination in a vaccine may be viable from a formulation standpoint. Since the Hib conjugate is polydispersed, comparing the amount of vaccine eluting using peak height alone, could have been a less reliable indicator had the Hib conjugate been from different batches or manufacturers. But since in this study, the Hib conjugate was from only one batch and manufacturer, measuring the height of a prominent peak alone was a useful indicator of the amount of vaccine eluted. Peak area which is usually a more reliable indicator, could not be used in the comparison because the broad elution time of the Hib-CRM197 overlapped with that of the MenC-CRM197 in their combination as seen in Figure 3. Generally, a combination of vaccines with identical conjugate proteins and same charge of polysaccharide subunits do not interact physico-chemically when in a suitable buffer of appropriate pH and sufficient ionic strength. As anionic polysaccharides would be expected to repel each other34, the polysialic acid of the MenC is not expected to bind to the phosphorylated PRP of the Hib, since both of them are negatively charged at around neutral pH. An NMR spectroscopic study of CRM197-conjugated meningococcal vaccines has found a high degree of flexibility of the conjugates in solution, and despite the highly anionic nature of serogroup C, W and Y conjugates, a tendency for the conjugates to aggregate at high concentration, although this NMR study was performed with conjugates in very low ionic strength (5 mM) phosphate buffer35. The effect of different buffers on the elution of the conjugates, and to a lesser extent, the protein markers, suggested that the vaccine-column matrix interactions were affected by the buffer salt. Phosphate ions bind proteins & metal cations with higher affinity than the `biological buffer' salts such as MOPS, HEPES and others 36. Different buffer ions can interact differently in solutions with anionic saccharides (conjugates) such as found in vaccines with group A, B, W135 and Y meningococcal capsular polysaccharides as well as many of the pneumococcal polysaccharides serotypes. This study did not however assess interactions between the tetanus toxoid (TT)-based conjugates, Hib-TT and MenC-TT, which are combined in the Hib/MenC booster vaccine licenced for use in the U.K. in 2005 by the Medicines and Healthcare Products Regulatory Agency. Further work has addressed the effect of carrier protein, saccharide type, buffer and pH in a study on aluminium adjuvant adsorption. Differences in aluminium adjuvant binding if combined with different adjuvants in combination vaccines would be expected, however, to occur. Investigation of the effect of the manufacturers' formulation buffers and diluents for reconstitution on vaccine interactions and adsorption to antigens and adjuvants should be determined as appropriate in physico-chemical and immunological studies in the laboratory, as clinical studies cannot ethically evaluate the feasibility of making new or tailor-made combinations. Since some combined vaccine preparations containing purified B. pertussis proteins and pertussis toxoid, such as DTaP-Hib, are known for inducing lower Hib antibody levels following routine vaccination5, 12, 13 and in laboratory models 37, physico-chemical interactions amongst different individual vaccine components should also be investigated15. Furthermore meningococcal serogroup polysaccharide-protein conjugate vaccines should be studied for possible interactions with Hib conjugate vaccines, since tailored polyvalent meningococcal polysaccharide are currently on the market and new conjugate vaccines are in the pipeline to meet the needs around the world 38, 39. Acknowledgements We are grateful to Dr. Stephen Harding, course director for the MSc program in Applied Biomolecular Technology at the University of Nottingham, to Dr. Michael J. Corbel, Head of the Division of Bacteriology, NIBSC, for facilitating this interesting and enjoyable project, and, to NIBSC and Divisional staff for their willingness to help. Much appreciation goes to Drs. Paolo Costantino and Aldo Giannozzi, Novartis Vaccines and Diagnostics, Siena, Italy for the generous gift of vaccine material and for fruitful collaborations. References
Copyright © 2007 - Makerere Medical School, Uganda The following images related to this document are available:Photo images[hs07038t1.jpg] [hs07038f3.jpg] [hs07038f4.jpg] [hs07038f2.jpg] [hs07038f1.jpg] [hs07038t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}