 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Health Sciences, Vol. 8, No. 1, March, 2008, pp. 25-35 Enhanced antimalarial effects of chloroquine by aqueous Vernonia amygdalina leaf extract in mice infected with chloroquine resistant and sensitive Plasmodium berghei strains Iwalokun, BA Department of Biochemistry, Lagos State University, Ojo, P.M.B 1087, Apapa -Lagos, Nigeria, E-mail: bamwal@yahoo.com Code Number: hs08007 Abstract Background: The emergence and spread of Plasmodium falciparum with resistance to chloroquine (CQ), the safest and
cheapest antimalarial drug coupled with the increasing cost of alternative drugs especially in developing countries have necessitated the need
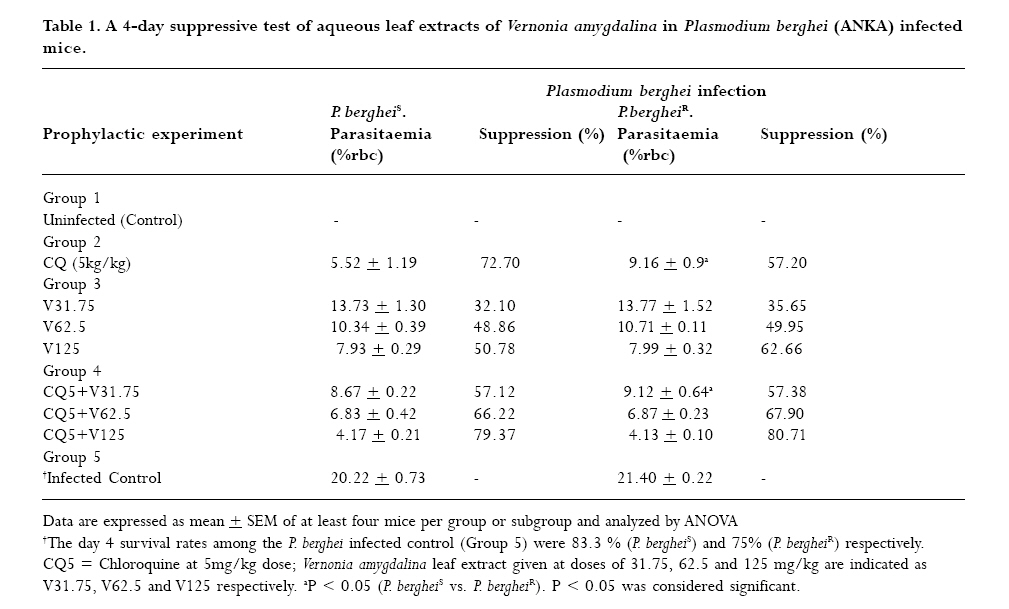
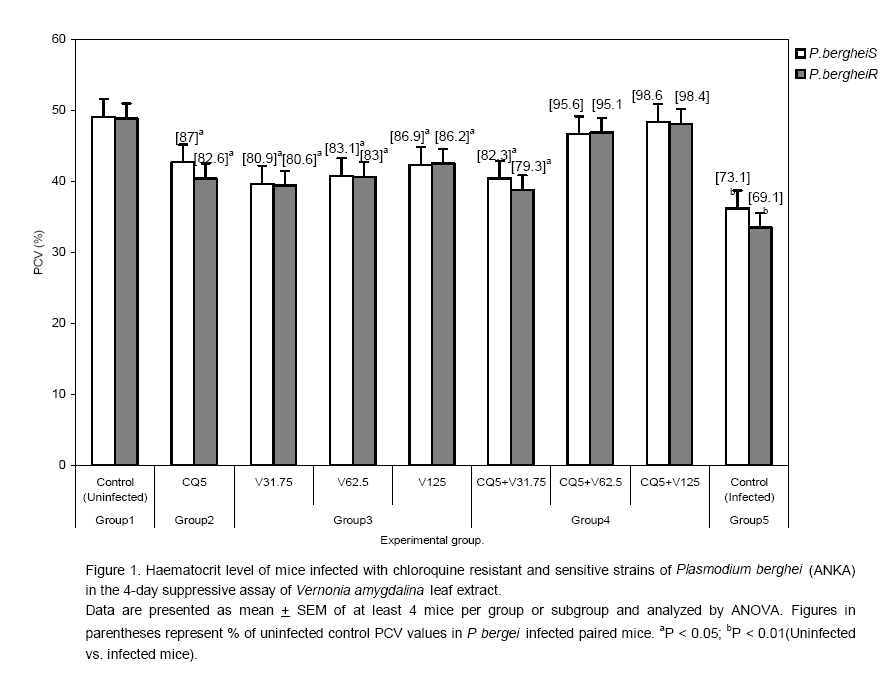
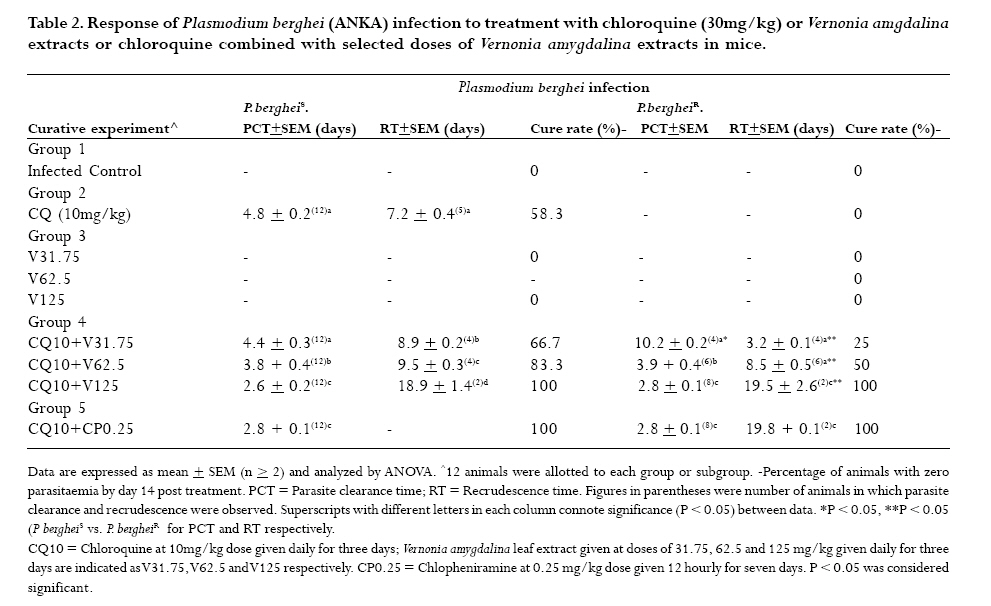
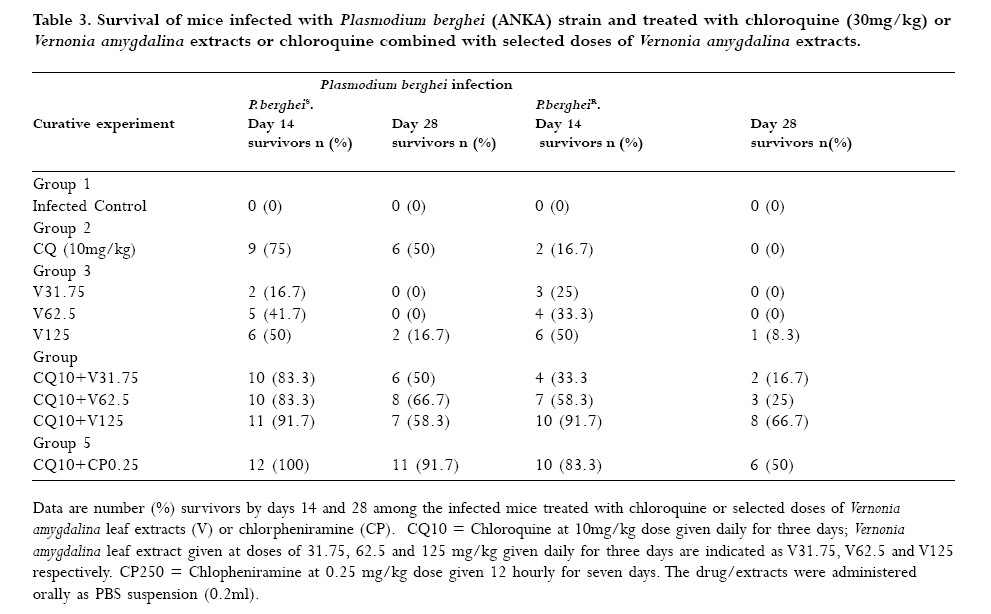
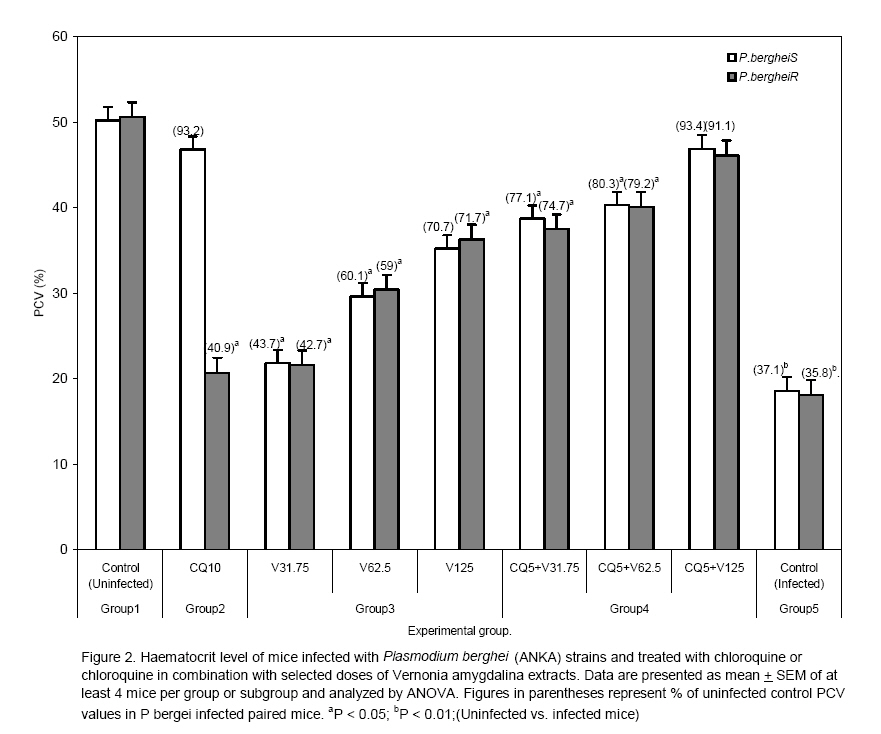
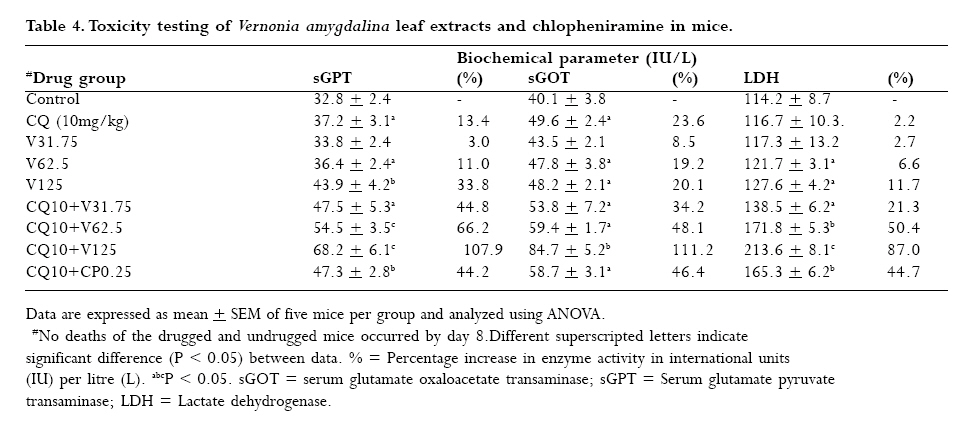
to optimize antimalarial actions of plant extracts and restore chloroquine efficacy. Key words: Plasmodium berghei infection, chloroquine resistance, vernonia amygdalina extracts, chloroquine -vernonia combination, Toxicity. Introduction Malaria remains the greatest cause of hospitalization and deaths among the parasitic infections that afflict man [1]. Over 40% of the world's population are at risk of malaria and active clinical cases due to Plasmodium falciparum parasitaemia is estimated to be 300 - 500 million annually with over 90% occurrence mostly in children and primigravidae in Sub-Saharan African. 1 -3% of these cases result in 1.5-2.7 million deaths due to anaemia, cerebral malaria and acidotic complications 2, 3. Chemotherapy remains the kernel of malaria control and the previously efficacious chloroquine has substantially failed as a prophylactic and therapeutic antimalarial in many endemic countries of Africa, Asia and South America due to a corresponding spread of chloroquine resistant P. falciparum strains with mutant alleles for chloroquine resistance transporter proteins (pfcrtT76) and multidrug resistance glycoprotein-1 (pfmdr-1Y86) 4,5. Consequently, the diminished potency of chloroquine above the 25% failure in many of the endemic countries including Nigeria, have paved way for the use of other antimalarial drugs in the armamentarium. Unfortunately, these alternative antimalarial drugs including sulphadoxine-pyrimethamine, mefloquine and halofantrine have also been observed to have their efficacy comprised by resistance developed by P. falciparum against them 6, 7. Furthermore, these drugs are not only costlier than chloroquine but also parade heterogenous levels of toxicity and may invoke poor compliance in patients 8. Artemisinins in combination with other antimalarials with distinct mode of actions remain the only regimens for which clinical failures have not been documented 9. However, most of these regimens are administered in loose doses and are currently having difficulty in implementation especially in rural areas of developing countries due to affordability and accessibility constraints 10, 11. Surprisingly, chloroqune remains the regimen of choice against malaria in pregnancy due to its enormous safety and cheapness, which remains insurmountable by other antimalarial alternatives 12. Taken together, the above findings have provided a justifiable evidence and need to develop strategies of restoring chloroqune efficacy and prolong its clinical utility as an antimalarial drug. In the last twenty years, these strategies have employed pharmaceutical drugs masquerading as antidepressants (e.g. fluoxetine and desipramine), antihistamines (e.g. promethazine and chlorpheniramine), calcium channels blockers (e.g. verapamil and nifedipine) and antipsychotics (e.g. chlorpromazine) to reverse chloroquine resistance by P. falciparum in vitro and in vivo in rodent and apes malaria models 13, 14, 15, 16, 17. In these studies, positive outcomes include reduced 50% inhibitory concentrations of chloroquine (IC50s), increased chloroquine uptake and chloroquine ferriprotophorphyrin IX binding 14, 17. However, the replication of these therapeutic benefits in human malaria has been reported to fail with desipramine 18 but achieve success with chlorpheniramine in Nigeria 16. Furthermore, apart from the obvious elevated cost of treating malaria, many of these chemosensitizers have multisystemic effects, may elicit side effects and their various doses of chloroquine resistance reversal can be toxic 19 . There are also strong indications that their use may be affected by behavioral factors in rural areas where herbal extracts are still used to treat febrile illnesses including malaria in these populations20 . This scenario has re-awakened interest in the exploration of medicinal plants from which antimalarials such as quinine and artemisinin were found 21. For instance the recent work of Tona et al 22 provided a scientific validity for the traditional use of eight medicinal plants including Vernonia amygdalina for the treatment of malaria in the Democratic Republic of Congo. Vernonia amygdalina is a perennial herb belonging to the Compositae family, extracts of the plant have been used in various folk medicines as a remedies against helminthic, protozoal and bacterial infections with subsequent scientific supports to these claims 23. The findings of Tona et al21, Abosi and Raseroka 24 and Massaba 25 indicate that Vernonia amygdalina leaf extract holds promise as an antimalarial agent despite its inability to clear parasitaemia completely. In this regard, the possibility of vernonia enhancing the efficacy of chloroquine needs to be explored. In the context of Complementary and alternative medicine, we hypothesize that the outcome of this study would support or question the feasibility of developing cost effective and efficacious chloroquine-based antimalarial therapies with herbs especially in developing countries. In the present study, various doses of aqueous vernonia leaf extract were combined with sub-curative and curative doses of chloroquine to investigate the effect of the plant on the prophylactic and chemotherapeutic efficacy of chloroquine in mice infected with chloroquine resistant and sensitive Plasmodium berghei strains. The efficacy of Vernonia amygdalina as a chloroquine resistance reversal agent was also compared with that of chlorpheniramine. Materials and methods Vernonia amygdalina (Compositae) Samples of Vernonia amygdalina plant were uprooted from the soil in a local farm situated less than 5 km from Lagos State University, Ojo, Nigeria and were taken to Forestry Research Institute, Ibadan Nigeria for authentication followed by deposition of the leaf as a herbarium collection with voucher number: FHI 106509. The leaves, being detached from the plant stems and sieved to remove debris were then taken to the Pharmacognosy Department of College of Medicine, University of Lagos, where they were oven -dried at 50°C for 5 days and ground to powdery form prior to extraction. Plant extraction 500g of ground Vernonia leaves were stepped in sterile water (1500ml) at 26°C for 6 days under sterile conditions. On day 7, the concoction was filter sterilized by passage through a 45?m Millipore membrane filter (St Quentine, France). The filtrate was then freeze- dried under low pressure (Memmer, Ltd, Germany). The freeze dried sample was subsequently reconstituted with sterile phosphate buffered saline (PBS, pH 7.2) to 31.75 -125mg/kg mouse body weight doses, designated as V31.75, V62.5 and V125 and stored in sterile bottles at 4°C until ready for use. Drugs Tablets of chloroquine phosphate, CQ (Pfizer, Nigeria) and chlopheniramine maleate, CP (Roche, Nigeria) were dissolved in PBS to final doses of 5mg/kg and 10mg/kg body weight for chloroquine (CQ) and 0.25mg/kg body weight for CP (CP 0.25). Animals Inbred male and female (non-pregnant) Swiss albino mice weighing between 19 and 22g (mean weight, 20.4g) were obtained from the Animal House of College of Medicine, University of Lagos. The animals were observed under 12h light/dark cycle in metabolic cages in a well ventilated rodent cubicle. They were fed a mice pellet diet (Ladokun Farms, Nigeria) with water ad libitum. The caring and experimental use of the mice was in accordance with National Institute of Health Guidelines for Care of Laboratory animals [26]. The animals were acclimatized for 7 days prior to their randomization into the various experimental groups. Parasite The two Plasmodium berghei clones used in this study were obtained from the Department of Biochemistry, Nigerian Institute of Medical Research, Lagos, Nigeria. The parasites are ANKA clones with chloroquine resistance (P. bergheiR) and sensitive (P. bergheiS) phenotypes and maintained by serial passage in mice with and without chloroquine pressure (5mg/kg). Parasite density determination and inoculum preparation A standard inoculum was prepared from a donor mouse with P. berghei parasitized erythrocytes. Infected blood from the donor mouse was obtained by cardiac puncture after anesthesia with chloroform Parasitaemia was established by microscopic examination of a thin blood film under oil immersion at x 1000 magnification and measured as a percentage of infected erythrocytes in fields of 500 erythrocytes. Each mouse was infected with a standard inoculum of the 106 parasitized erythrocyte suspension in phosphate buffered saline (0.2 ml) from a donor mouse that was prepared based on percentage parasitaemia and number of erythrocytes counted per microlitre of blood using an improved Neubear Haematocytometer. Prophylactic treatment Here, a 4-day suppressive test according to Peter et al[27] was used. The animals were divided into 5 groups of 12 mice each. Animals in group 1 were given the vehicle only (i.e. 0.2ml PBS) and served as the uninfected control. Animals in group 2 were administered 5mgkg suppressive dose of chloroquine and those in group 3 were further divided into three subgroups and administered vernonia extracts at 31.75, 62.5 and 125mg/kg doses. In group 4, the experimental animals were also divided into three subgroups and administered chloroquine in combination with the three selected do ses of vernonia extracts. Animals in group 5 (infected control) were also given the vehicle, but randomized into two subgroups before chloroquine resistant and sensitive parasite challenge. Each mouse was intraperitoneally infected with 106 parasitized erythrocytes in 0.2 ml PBS suspension. Treatments commenced immediately after inoculation (Day 0) and continued on days 1, 2 and 3 via an oral route using a canula. Tail blood films of the infected animals were prepared, fixed in methanol, stained with 4% Giemsa stain for 20 min and examined microscopically under oil immersion on day 4 post inoculations. Haematocrit (PCV) reading for each animal was also taken after centrifugation of whole blood in sealed heparinized capillary tube [28] on the day of parasitaemia evaluation. Parasite density and percentage reduction in parasitaemia relative to chloroquine control on group-by-group basis were calculated. Curative treatment In this experiment, another set of 60 albino mice randomized into 4 groups of 12 animals per group were intraperitoneally infected with 106 parasitized erythrocytes with oral treatments commencing on day 3 post inoculation. Animals in group 1 were given the vehicle only and served as the control. In group 2, the animals were given a curative dose (10mg/kg) of chloroquine daily for 3 days. Animals in group 3 were further divided into three subgroups and administered the selected 31.75mg/kg, 62.5mg/kg, and 125mg/kg doses of vernonia extracts. In group 4, the mice were also randomized into three subgroups and treated with chloroquine (10mg/kg) in combination with 31.75mg/kg, 62.5mg/kg, and 125mg/kg of vernonia extracts. Animals in Group 5 were treated with CQ (10mg/kg) in combination with 0.25 mg/kg of chlorpheniramine maleate. The chemosynthesizer was administered 12 hourly for seven days. Tail blood films were prepared starting from day 3 post inoculation on a daily basis till day 9 and thereafter every other day till day 28. Parasite clearance time (PCT) defined as time in hours to disappearance of peripheral parasitaemia in the infected mice following treatment and recrudescence time (RT) defined as time to reappearance of parasitaemia during the follow-up period were determined in all the treated groups. The number and percentage of survivors by days 14 and 28 were also recorded. Level of parasitaemia was also determined as previously described. Plasma preparation and toxicity testing. Here, the animals were orally administered with 0.2ml PBS (control), 10mg/kg chloroquine, V31.75, V62.5, V125, and 0.25.mg/kg CP every 24 hours for 7 days. On day 8, the animals were lightly anaesthetized with chloroform and subsequently sacrificed by cervical decapitation. Whole blood was collected into EDTA test tubes, mixed by rolling and then centrifuged at 5000 rpm for 5 min. The resulting plasma was subsequently employed for liver enzyme assays. The enzymes assayed for were glutamate -pyruvate (GPT) and glutamate -oxaloacetate (GOT) transaminases according to Bergmeyer and Bent [29] and lactate dehydrogenase as described by Wroblewski and La Due [30]. Statistical analysis Data expressed as mean ± SEM was analyzed for significance of disparity using a 2-way analysis of variance (ANOVA) and the F-distribution. P < 0.05 was regarded as significant. Results The data on day- 4 parasitaemia suppression by chloroquine (5mg/kg) and selected doses (31.75 - 125mg/kg) of Vernonia amygdalina leaf extracts is presented in Table 1. In the absence of prophylactic intervention, highest parasitaemia due to chloroquine sensitive (20.22 ±0.73 %rbc) and resistant (21.40 ± 0.22 %rbc) Plasmodium berghei infection were observed in the infected control (Group 5). However, chloroquine elicited suppression of parasitaemia by 72.70% and 57.20% respectively in the animals infected with P. bergheiS and P. bergheiR. The disparity in the resulting day 4 parasitaemia (5.52 ± 1.19 vs. 9.16 ± 0.9 % rbc was found to be significant (P < 0.05). Prophylactic control of parasitaemia with the selected doses of Vernionia amygdalina extracts 31.25 -125mg/kg) also revealed dose-dependent suppression of parasitaemia of comparable magnitude (32.10-79.37% vs. 35.65-80.71%) in P.bergheiS and P. bergheiR infected mice when administered singly or in combination with chloroquine. Prophylactic blood haematocrit level was found to reduce significantly (P < 0.05) to 79.3 -87% of uninfected control in infected animals treated with chloroquine, 31.75 -125mg/kg extracts alone and combination with chloroquine at 31.75mg/kg dose (Figure 1). The extracts were also tested for their ability to clear parasitaemia due to P. bergheiS and P. bergheiR. Unlike chloroquine at curative dose of 10mg/kg, which produced clearance time to 4.8 ± 0.2 days and day 14 cure rate of 58.3% in mice with P. bergheiS malaria, data indicate 100% failure rate (cure rate = 0%) when the extracts were used singly but dose-dependent cure rate of 66.7 -100% when used in combination with chloroquine (Table 2). Chloroquine-chlorpheniramine combination also cleared parasitaemia (PCT = 2.8 ± .0.1 days) and elicited 100% cure rate by day 14 in both P. bergheiS and P. bergheiR infected malaria suggesting reversal of chloro quine resistance in the latter. However, the P. bergheiR parasites recrudesced with recrudescence time of 19.8 ± 0.1 days that was comparable to 18.9 ± 1.4 and 19.5 ± 2.6 days produced by chloroquine-V125 combination in P. bergheiS and P. bergheiR infected mice (Table 2). The extracts were also tested for their ability to clear parasitaemia due to P. bergheiS and P. bergheiR. Unlike chloroquine at curative dose of 10mg/kg, which produced clearance time to 4.8 ± 0.2 days and day 14 cure rate of 58.3% in mice with P. bergheiS malaria, data indicate 100% failure rate (cure rate = 0%) when the extracts were used singly but dose-dependent cure rate of 66.7 -100% when used in combination with chloroquine (Table 2). Chloroquine-chlorpheniramine combination also cleared parasitaemia (PCT = 2.8 ± .0.1 days) and elicited 100% cure rate by day 14 in both P. bergheiS and P. bergheiR infected malaria suggesting reversal of chloroquine resistance in the latter. However, the P. bergheiR parasites recrudesced with recrudescence time of 19.8 ± 0.1 days that was comparable to 18.9 ± 1.4 and 19.5 ± 2.6 days produced by chloroquine-V125 combination in P. bergheiS and P. bergheiR infected mice (Table 2). Generally, parasite clearance time and recrudescent time were found to be reduced and prolonged significantly with increased doses of Vernonia amygdalina extracts in combination with chloroquine irrespective of the resistance status of the parasite (Table 2). The data presented in Table 3 shows dose-dependent percentage survival (16.7 -50%) in mice with P. bergheiS malaria treated with the extracts alone by day 14 post treatment. By 28 post treatment, survivors (8.3 -16.7%) were found only in the V125 treatment arm in both P. berghei infected groups. While treatment of the infected mice with chl;oroquine (10mg/kg) alone produced 6 (50%) survivors by day 28 only in the P. bergheis, combination treatment with selected doses of Vernonia amygdalina produced an improvement in survival rate in these animals with CQ-V62.5 and CQ-V125 eliciting highest survival rates of 66.7% each in the treated P. bergheiS and P. bergheiR infected mice (Table 3). Compared to the uninfected control, the P. berheiR-infected mice treated with a curative dose of chloroquine (10mg/kg) were also found to elicit a significant (P < 0.05) decrease in haematocrit level (40.9 vs. 100%) but which improved with increased doses of the combinational Vernonia amygdalina extract achieving a non-significant level in the CQ-V125 treatment arm in both P. bergheiS and P. bergheiR infected mice groups (Figure 2). Table 4 shows the mean activity of sGPT, sGOT, and LDH in health mice administered via the oral route with the selected doses of Vernonia amygdalina leaf extracts alone of in combination with chloroquine (10mg/kg) for 7 days. Compared to the control (undrugged mice) with mean activity of 32.4 ± 2.4 IU/L for sGPT, 40.1 ± 3.8 IU/L for sGOT and 114.2 ± 8.7 IU/L for LDH, chloroquine alone was observed to cause elevation in transaminase and LDH activity by 13.4 -23.6% and 5.6% respectively. A similar but dose-dependent increases in sGOT (3.0 -33.8%), sGPT (8.5 -20.1%) and LDH (2.7 -11.7%) were further observed in mice drugged with the extract alone. Vernonia -chloroquine combination drugging resulted in further elevation in these parameters with CQ + V125 combination eliciting increases greater than those of CQ + CP 0.25 (87.0 -111.2 vs. 44.2 -46.4%; P < 0.05). However, no deaths of drugged and undrugged mice occurred by day 8 of analysis. Discussion The increasing failure of antimalarias including chloroquine and sulphadoxine—pyrimethamine in many endemic sites of Africa has generated tremendous public health concern necessitating for more exploration of plants for newer antimalarial agents and optimization of those with existing antimalarial activity when used singly, as composite remedies or in combination with orthodox medicine31. The present study has not only validated the antiplasmodic activity of Vernonia amygdalina but has also demonstrated the inherent ability of the plant's aqueous leaf extract to work in favour of the restoration of chloroquine efficacy as a prophylactic and chemotherapeutic agent against chloroquine sensitive and resistant P. berghei infection in mice in vivo. This is because in the 4 -day suppressive assay, co-administration of vernonia with prophylactic dose of CQ resulted in the dose-dependent suppression of parasitaemia that was greater than the effects of chloroquine or selected doses of the plant extracts when used singly in P. bergheiR infected mice. These combinations particularly CQ + V125 also suppress parasitaemia due to P berghei with comparable prophylactic efficacy with chloroquine (5mg/kg) when used singly. The outcome with chloroquine was not unexpected since the parasite had a sensitive phenotype. A similar work by Okonkon et al 32 also revealed the efficacy of Hippocrata africana and Uvaria chamae but at larger doses of 200 -900mg/kg, Garavito et al [33] had previously reported the alkaloid fraction of Abuta grandifolia to elicit 66% suppression of P. berghei parasitaemia at 250mg/kg. While in the work of Okpako and Ajaiyeoba34, the prophylactic functions of Striga hemothica and Tapinanthus sessilifolius extracts characterized by 51 -68% suppression of P. berghei parasitaemia in mice was reported. Aside their high dose-anti-berghei efficacy, these plants were tested uncombined with chloroquine and also noted for toxicity [35]. Meanwhile, a few recent studies have reported antimalarial efficacy against P. falciparum and P. berghei of some singly applied plant extracts at microgram/kg doses 19. But these observations were made in vitro based on parasite cultures, which are often influenced by culture medium and serum factors 36. However, in terms of orthodox medicine -plant extract combination, a few studies have also been reported. They include CQ-Ekerbergia capensis and CQ-Clerodendrum myricoides efficacy against multidrug resistant P. falciparum isolates 37 in culture, CQ-Allium sativum alone against P. berghei malaria38 and Azadiracta indica with pyrimethamine against P. yoeli nigeriensis malaria in mice 39. These combinations except Azadiracta indica -pyrimethamnine at 800mg/kg [39] were found to lack prophylactic efficacy. A prophylactic study by Agbaje and Onabanjo 40 however revealed 96-hour protection against P. yoeli infection in mice by ethanolic extract of Enanthia chloranthia at £ 150mg/kg. Therefore, our observation that Vernonia amygdalina at 125mg/kg in combination with CQ suppress parasitaemia by more than 70% on day 4 (96 h) connotes a stronger prophylactic action. Dose-dependent suppression of heamatocrit reduction due to parasitaemia was also observed; indicating that of Vernonia may inhibit or compensate for erythrocyte destruction due to parasitaemia. Agomo et al 41 had previously reported an increased reticulocyte counts in P. yoelii nigeriensis infected mice treated with Azadiracta indica, Morinda lucida and Enantia cloranthia extracts to support this possibility. Moreso that greater improvement was observed in the therapeutic experiment. That the efficacy of an antimalarial drug can be enhanced by combination with plant extracts has been found with Azadiracta indica-chloroquine combination at 800mg/kg 39 in P. yoelii malaria ,Tinospora cordifolia-chloroquine combination in P. falciparum malaria via a daily 300mg dosing42, E. capensis and C. myricoides in combination with CQ with IC50 < 30 microgram/kg 37 and Khaya grandifolia-CQ against P. falciparum with IC50 < 9.63microgram 43 (Bickii et al, 2000) and guanghuoxiang-sodium artesunate combination with IC50 < 100 microgram/kg44. It is of interest to know the higher therapeutic efficacy observed in some of these combinations are due to the active Phytoconstituents in the plant and not their crude preparations as tested in this study. For instance, limonoids and gedunin were responsible for the potency of Khaya grandifolia combination [43]. While it is the essential oil of guanghuaxiang that was tested in the combination with sodium artesunate [44]. While unlike the present study, in vitro culture assays were also used in these studies to monitor anti-plasmodial activity of the various combinations tested [37, 43, 44]. Meanwhile, other anti-plasmodial phytoconstituents reported include quinoline alkaloids such as cephantharin, cryptolepine, isocryptolepine and neocryptolepine, courmarins and terpenoids[45, 46, 47, 48] The last two Phytoconstituents are abundantly present in Vernonia sp. and may thus contribute to the observed anti-berghei properties of the plant. However, the 62.5mg/kg and 125mg/kg combination doses at which Vernonia was found therapeutically effective in this study is relatively lower than effective doses reported by Nwafor et al [39] for Azadiracta indica (800mg/kg), and those previously reported for Vernonia amygdalina leaf extract [24, 25]. Plant extracts of Bidens pilosa, Cochlospermum planchonii have also been found to elicit antiplasmodial activity but with lower than 60% parasitaemia reduction profile at day 7 posttreatment[49, 50] . A randomized controlled trial study by Mueller et al [51] using Artemisia annua extract as antimalarial agent also revealed 74% cure rate for the plant but with high recrudescent rate thereby withdrawing its recommendation as an alternative to modern antimalarials. In this study, we observed Vernonia in combination with chloroquine to dose dependently prolonged recrudescence in < 50% of both the chloroquine sensitive and resistant P. berghei infected mice. This observation further reveals an anti-recrudescent potential of Vernonia amygdalina especially when administered at 125mg/kg dose. Moreso that these combinations also produced anti-berghei effects comparable to chlorpheniramine-CQ in terms of parasite clearance time (PCT) and recrudescent time (RT) particularly in the experimental chloroquine resistance Plasmodium berghei infection in the mice. Chloroquine at therapeutic concentration in the parasite food vacuole, functions as a schizonticidal agent via arrest of DNA replication, inhibition of hemozoin formation and energy deprivation via interactions with parasite lactate dehydrogenase2, 53 This chloroquine actions are prevented in resistance strains owing to parasite mediated alkalinisation and efflux of the drug from the food vacuole 53 but are restored by chemosensitizers such as chlorpheniramine as observed in this study. Therefore, the ability of Vernonia amygdalina leaf extract to elicit similar activity as chlropheniramine is suggestive of the chemosensitizing (i.e. chloroquine resistance reversal) potential of the plant when used as an adjuvant with chloroquine. The action of vernonia may also involve prolongation of elimination half life chloroquine as reported for Azadiracta indica 39 or synergistic action with chloroquine in inhibiting hematin formation as reported for Humulus lupus 24, 25. Therefore, there is a need to advance the current status of Vernonia amygdalina to an antimalarial leas drug level through isolation and characterization active antiplasmodial components in the extracts, known to contain many terpenoid and alkaloid compounds. The possibility of safety following the use vernonia at combinational doses of 62.5 and 125mg/kg can be explained by the observed moderate - mild elevations in the plasma levels of liver enzymes in the experimental animals, which chlorpheniramine used presently in the management of uncomplicated malaria in Nigerian children [16] also elicited. In vitro cytotoxic effects of chlorpheniramine against human cell lines and cancer cells have been reported in a few studies 55. Taken together, the present study has demonstrated the potential of vernonia to reverse chloroquine resistance when used as an adjuvant with chloroquine to treat malaria. Further development of Vernonia for this purpose would no doubt provide cost effective strategy of malaria control especially in developing countries where poor affordability and accessibility to orthodox drugs are the primary barriers to health care. Further studies involving the use of Plasmodium falciparum strains and their in vitro culture coupled with toxicity testing for the assessment of drug-extract efficacy and safety are being carried out. Acknowledgement The technical assistance rendered by staff in the animal care unit of Nigeria Institute of Medical Research (NIMR), Lagos-Nigeria is sincerely appreciated. References
Copyright © 2008 - Makerere Medical School, Uganda The following images related to this document are available:Photo images[hs08007t1.jpg] [hs08007f1.jpg] [hs08007t4.jpg] [hs08007f2.jpg] [hs08007t3.jpg] [hs08007t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}