 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Health Sciences, Vol. 9, No. 2, June, 2009, pp. 66-74 Cytokine profiles and antibody responses to Plasmodium falciparum malaria infection in individuals living in Ibadan, southwest Nigeria *ô,§ Nnaemeka C. Iriemenam, §,¶,‡ Christian M. F. Okafor, §Halima A. Balogun, ÆIdowu Ayede, ¶,††Yusuf Omosun, †Jan-Olov Persson, §Margareta Hagstedt, ¶Chiaka I. Anumudu, ¶Roseangela I. Nwuba, §Marita Troye-Blomberg , §Klavs Berzins ôTropical Diseases Research Laboratory, Department of Medical Microbiology & Parasitology, College of Medicine of the
University of Lagos, Idi-araba, P.M.B. 12003 Lagos, Nigeria Code Number: hs09014 Abstract Background: The ability of the host immune system to efficiently clear Plasmodium falciparum parasites during a malaria
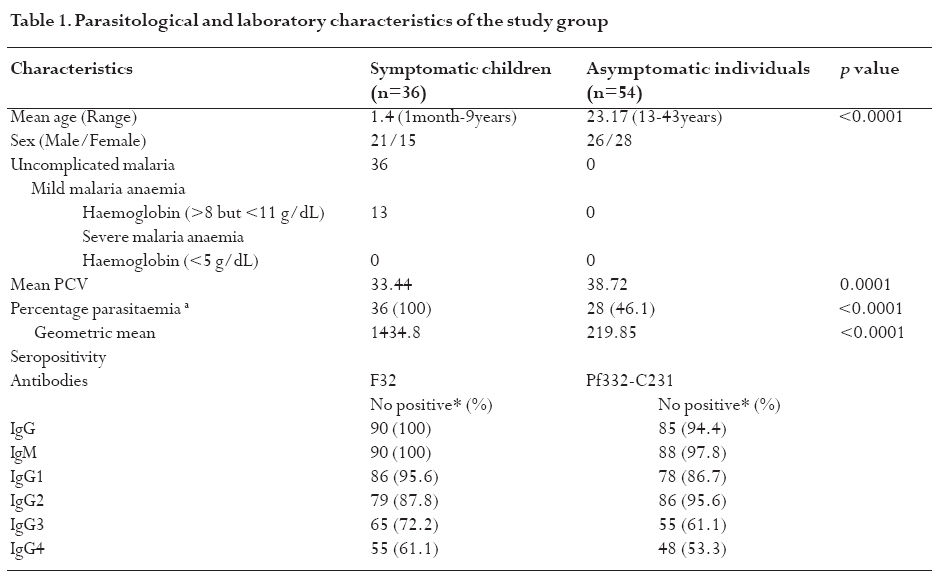
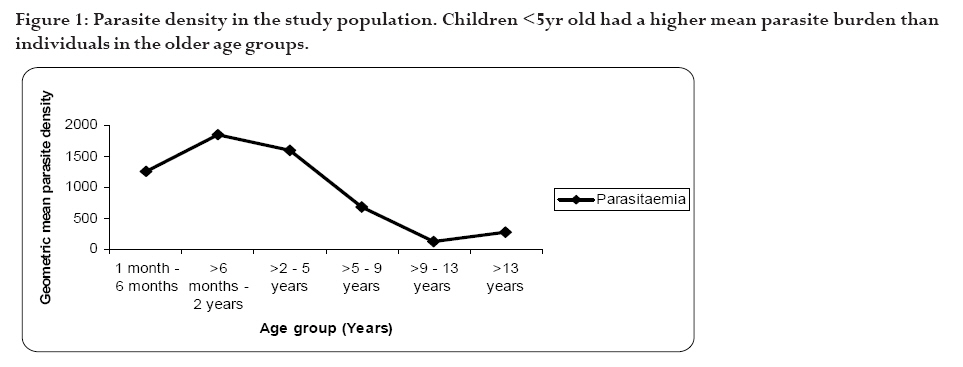
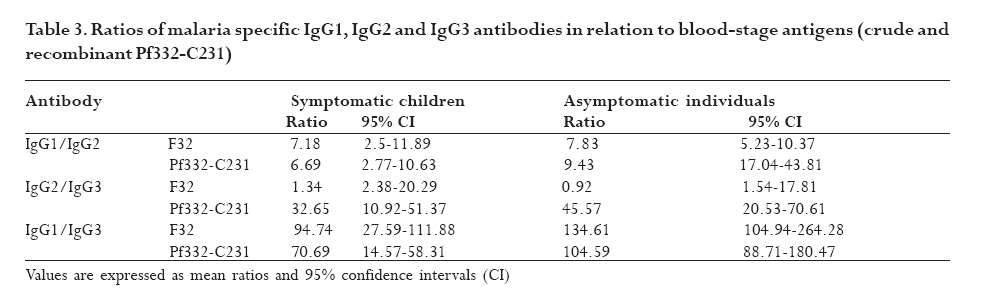
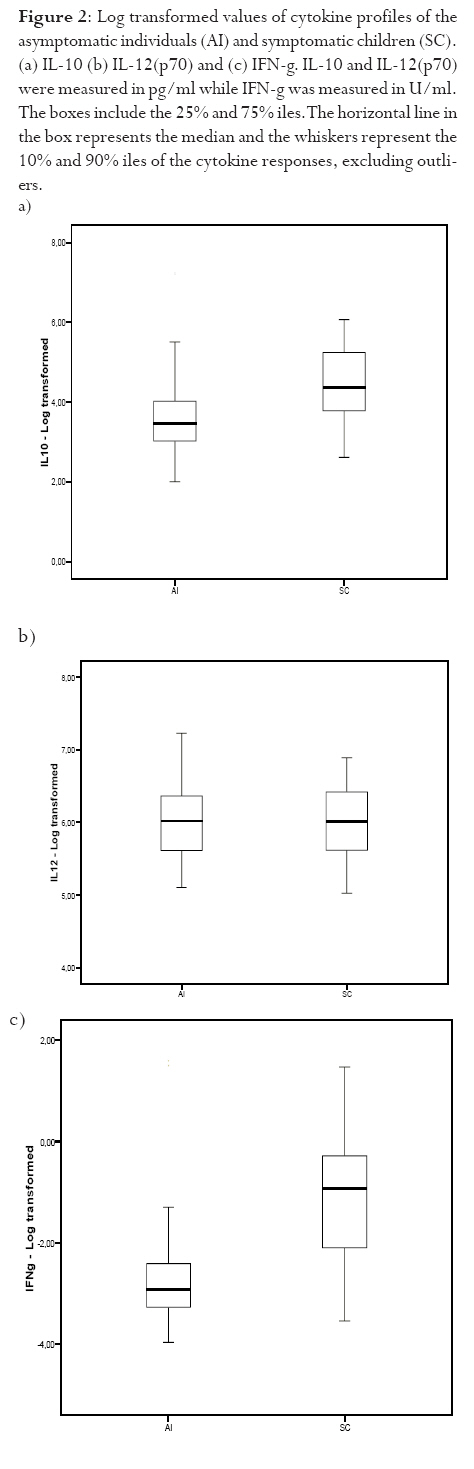
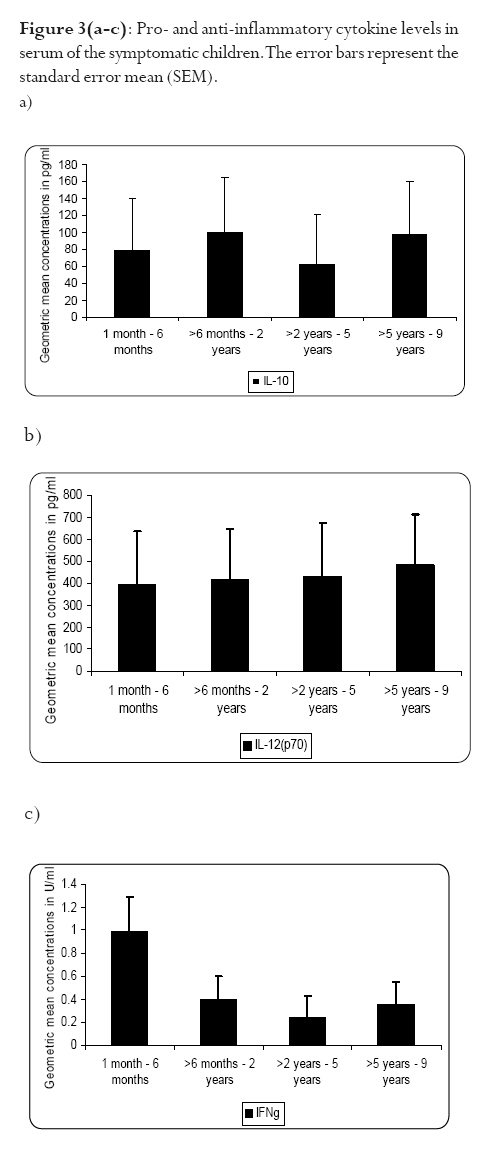
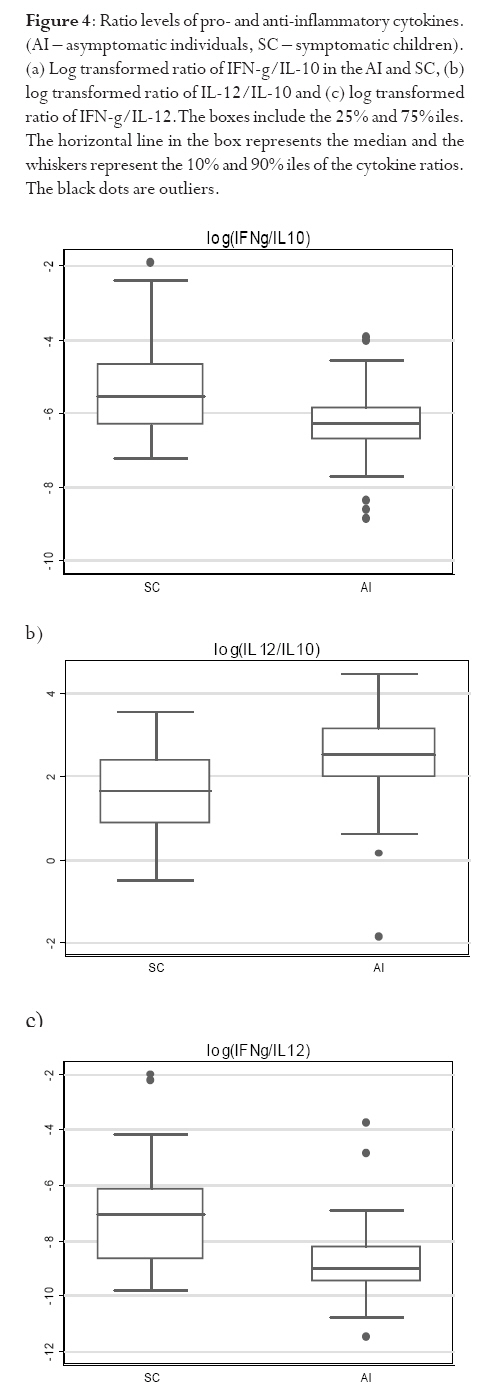
infection depends on the type of immune response mounted by the host. Keywords: malaria, antibodies, cytokines, anaemia Running title: Differences in T-cell and antibody responses to malaria Introduction Malaria remains one of the leading global health concerns with over 300 million clinical cases and more than 1 million deaths on annual basis1. Being a predominantly tropical disease, malaria is one of the top three killers among communicable diseases in Africa2. Non-immune individuals, pregnant women and children bear most of the morbidity and mortality due to the disease, which manifest as severe malaria anaemia and cerebral malaria complications. Severe falciparum malaria in children living in holoendemic endemic regions is characterized by high-density parasitaemia and severe malarial anaemia but less frequently cerebral malaria. Although the exact mechanisms underlying the pathogenesis of severe malaria are not well known, available evidence suggests that the pathology of the disease in humans is due to host immune responses against the parasite much more than the direct effect of the parasite3,4. Age and exposure to malaria infection are essential to the development of natural immunity to P. falciparum malaria. The need for better understanding of immunological basis of protective immunity to malaria is evident with the growing number of anti-malarial multi-drug resistance coupled with the vector's resistance to available insecticides. Both antibody dependent and cell-mediated mechanisms contribute to immune protection against the asexual blood stages of the parasite. The host immune responses against most infectious pathogens depend principally on the development of an adaptive immune response mediated by the release of cytokines produced by the appropriate T helper cells5. Th1 immune responses activate macrophages as part of cell-mediated immunity required for clearance of intracellular pathogens, whereas Th2 immune responses regulate humoral immune responses, stimulate growth of mast cells and also act to suppress cell-mediated immunity5,6. As a result, Th1 and Th2 cytokines work antagonistically to regulate each other's activities5. Blood-stage immunity in experimental models and in humans is dependent on CD4+ T cells subset, B cells and antibodies while the CD8+ T-cell subset has been associated with cytolytic activity against the parasite in liver stages7. The secretion of antibodies is a significant part of the host immune response to malaria infection; these antibodies are of different isotypic specificities, thereby exerting different functional capacities. The fact that IgG1 and IgG3 antibodies appear to be highest in protected individuals, supports their functional relevance in parasite neutralisation and opsonisation8. However, elevated levels of IgG2 antibodies with low levels of IgG4 in combination with allelic variants of FcgRIIa have also been associated with decreased risk of P. falciparum infection9. Pro-inflammatory cytokines have been associated with protective cell-mediated immunity by their capacity to induce parasite killing by monocytes/macrophages and neutrophils10. Anti-inflammatory cytokines counteract the production and possible cytopathic effects of pro-inflammatory cytokines10 and may thus be associated with malaria susceptibility11. Not much is known about the role of cytokines in regulating the immune response to malaria, but the relative balance between Th1 and Th2 cytokines is thought to be a crucial determinant of whether a response will be protective or pathologic12. We studied the cellular and humoral responses to P. falciparum parasite in individuals living in Ibadan, southwest of Nigeria to understand the responses that contribute to protective immunity. Materials and MethodsStudy area and individual enrolment We enrolled 36 children (<9years) presenting with malaria symptoms and 54 asymptomatic individuals (13-43 years) in a cross-sectional study in Ibadan, Nigeria (in 2005), a region of hyperendemic malaria transmission. The symptomatic children in this study were categorized into 2 groups (uncomplicated and severe malaria) based on the criteria given by the World Health Organisation (WHO)1,13. An individual was defined to have malaria if he/she complained of fever, had body temperature measured with an oral probe exceeding 37.5oC with asexual malaria parasites detected in the blood. Uncomplicated malaria was defined as P. falciparum parasitaemia of 1, 000 to 50, 000 parasites/ml, with glycaemia of >50 mg/dl and without severe malaria symptoms while severe malaria was defined with unarousable come (cerebral malaria), which persisted for at least 30 min after a seizure with severe malaria anaemia. Mild malaria anaemia was defined with haemoglobin (Hb) >8 but <11 g/dL in the presence of microscopically detectable asexual parasitaemia, while severe malaria anaemia was defined as Hb <5 g/dL or haematocrit <15%, with P. falciparum parasitaemia of >250,000 parasites/ml. None of the individuals had severe malaria at the time of the study. Thirteen individuals (10 males, 3 females) had mild malaria anaemia, and the rest (11 males, 12 females) had uncomplicated malaria. Samples were collected prior to any anti-malarial treatment. Anti-malarial administration with appropriate supportive therapy was based on the Nigerian Ministry of Health treatment policy for malaria in children. The study was approved by the Joint Ethical Committee of the College of Medicine and the University College Hospital, Ibadan, Nigeria. Sample collection Sample collection was done in the high transmission season. Informed consent was obtained from the parents/guardians of the children and from the adult donors before sample collection. Giemsa stained thick blood smears were done to determine the parasitaemia level and packed cell volume (PCV) were measured to determine haemoglobin levels from finger prick for each participant. Venous blood (3 ml) was collected from symptomatic individuals and 5 ml from the asymptomatic children using the heparinised vaccutainer system. Samples were transported at room temperature within 24hrs to the Department of Immunology, Stockholm University, Sweden where serum were separated immediately and frozen in -20oC until use. Serum for antibody determination was stored at 4oC in 1:10 dilution with Tris Hank containing sodium azide. Parasite cultures and antigen preparation F32 laboratory strain of P. falciparum was cultured under sterile conditions using Jensen candle jars as described by Trager and Jensen14. The parasites were maintained in in vitro culture medium with RPMI 1640 supplemented with L-glutamine (GIBCO, Grand Island, NY, USA), 1M Hepes, 7.5% NaHCO3, gentamycin 50 mg/ml, 200mM hypoxanthine and 0.5% albumax. The late stage infected erythrocytes were enriched using 60% percoll gradient separation with subsequent sonication to obtain the crude parasite antigen preparation15. Antibody and cytokine ELISA. P. falciparum blood stage crude malaria extract and C-terminal fragment of recombinant Pf332 (Pf332-C231) were used to determine antibody titres at 10 mg/ml and 5 mg/ml respectively in a standard indirect enzyme-linked immunosorbent assay (ELISA) procedures as previously described16. Serum pools of samples collected previously from clinically immune adults from Burkina Faso were used as positive control. The cut-off seropositivity values for antibody reactivity to the crude and Pf332-C231 antigens were the sum of mean antibody concentrations and 2SD of the values obtained with plasma from 10 non-malaria-exposed Swedish donors without any known prior exposure to P. falciparum parasite. Log-log correlation coefficients of the antibody standard curve were plotted and the results expressed as mean antibody concentrations. Detection of interferon gamma (IFN-γ), interleukin (IL-10) and IL-12p70 (all from Mabtech, Nacka, Sweden) in the serum were done according to the manufacturers' instructions. All samples were tested in triplicates. The optical densities (OD) were measured using the Molecular devices (VmaxTM Kinetic microplate reader, Menlo Park, CA, USA) at l405 nm. The detection limits were set according to the log-log correlative coefficient of the standard curve and also the background values of buffer and negative controls: IL-10 (10-10,000 pg/ml), IL-12(p70) (10-10,000 pg/ml) and IFN-γ (0.03-30 U/ml). Statistical analysis Analysis was done using statistical package Stata version 8 (College Station, Texas, USA) and SPSS version 15.0 for Windows (SPSS Inc., Chicago, USA). Data was normalised by log transformation. Univariate analysis of baseline variables between groups was performed using an unpaired Student's t test. The two-sample Wilcoxon rank-sum (Mann-Whitney U) test was used to compare variables within the groups. Coefficients of correlation with variables and parasitaemia were calculated by the Spearman's rho test. Logistic regression analysis was used to analyse the relationship between clinical malaria manifestations with antibody and cytokine responses adjusted for sex, age, anti-malarial antibodies and cytokine production respectively. A p value of < 0.05 was considered statistical significant. Results Parasitaemia and antibody response to antigens In this study, all the individuals that presented with malaria symptoms were positive for P. falciparum parasitaemia, but 46.1% of the asymptomatic individuals had Plasmodium parasitaemia. Mean parasite load was significantly higher (>6-fold) in the symptomatic children than their parasite positive-asymptomatic counterparts (Table 1). However, the mean age of the asymptomatic individuals is significantly higher than the symptomatic children. Parasite burden was highest in children less than 5years and subsequently reduced after that period (Figure 1). Peak parasite density occurred around 2years of age. Over 88% of the individuals in this study were sero-positive for IgM, IgG1 and IgG2 antibodies against either the crude or the recombinant antigen, comparatively fewer individuals were positive for IgG3 and IgG4. Effect of age on antibody levels Analysis of the relationship between age and antibody titres in individuals in this study showed that mean antibody levels were higher in the older individuals than younger children (Table 2). Stronger IgG1 and IgG3 subclass reactivity with crude malaria antigen was a general trend while there was a bias of IgG1 and IgG2 antibodies reactivity towards Pf332-C231 in the >5 years of age. The mean total IgG titres indicated a gradual decrease within the first two years of life but the levels increased after this period. Children <5 years expressed more IgM than IgG antibodies. IgM responses, however, were more pronounced in the symptomatic children <5years old and this same group had the highest mean parasite density (Figure 1). The levels of immunoglobulin isotypes IgG1, IgG2, IgG3 and IgG4 gradually increased with age and the highest levels were observed with IgG1 while IgG3 was notably low especially in the symptomatic children less than 5years old (Table 2). The levels of antibodies in individuals above 5 years were statistically significantly higher than the <5 years, except that of IgG4 antibody reactivity with crude malaria antigen. Comparison of the pattern of the IgG subclass antibody responses in relation to the blood-stage antigens was performed by calculating the ratios IgG1/IgG2, IgG2/IgG3 and IgG1/IgG3 (Table 3). IgG4 was not done due to its very low levels. Generally, the asymptomatic individuals had higher mean ratios with a strong reactivity towards IgG1 and IgG3 for crude malaria antigen and IgG1, IgG2 and IgG3 for the recombinant Pf332-C231 (Table 3). Logistic regression analyses showed no statistically significant differences between uncomplicated malaria and mild malaria anaemia children with reference to antibody levels responses adjusted for sex, age as confounding variables in the analysis. This was evident with odds ratio (OR) [95% confidence interval (CI)] using clinical malaria outcome as the dependant variable in the logistic regression analysis. However, there was a statistical significance with low levels of anti-Pf332-C231 IgG3 antibody in relation to mild malaria anaemia, which showed a confounding effect after adjustment for sex and age instead of an independent effect OR 0.098 [95% CI (0.005-1.14)] p=0.054). Association between parasitaemia and cytokine responses To establish differences in the degree of cytokine responses between symptomatic children and asymptomatic individuals, data for all individuals were compared. IL-10, IL-12(p70) and IFN-γ were detected in the serum from 86 (95.6%), 89 (98.9%), and 84 (93.3%) participants respectively. Distinct differences were observed between the cytokine responses seen in symptomatic children and the asymptomatic individuals (Figure 2). Analyses showed that the levels of IL-10 and IFN-γ in serum were significantly higher in symptomatic children than the asymptomatic individuals (p=0.025 and p=0.009 respectively). However, correlation analyses between cytokines and parasitaemia in symptomatic children revealed a good correlation with only IFN-γ responses with uncomplicated malaria R=0.93 and p<0.0001, but indicated a poor model and biased after adjustment R2=0.52. Furthermore, correlation analyses with IL-10, IL-12 and IFN-γ in the 28 asymptomatic individuals with parasitaemia revealed no statistical significance (R=0.18 p=0.2058, R=-0.18 p=0.1909 and R=0.21 p=0.1320 respectively) To determine whether the high levels of IL-10 and IFN-γ observed in the symptomatic children were restricted to P. falciparum malaria, we looked at the levels of each cytokine within each age group. The levels of IL-10 and IL-12(p70) were similar in all age-groups of the children (Figure 3a and b) whereas IFN-γ levels were significantly elevated within 1-6 months (Figure 3c) and then decreased across the groups. Using clinical malaria outcome as the dependent variable in the logistic regression model to analyze cytokine responses, we observed that IL-10 and IFN-γ levels were significantly elevated in 13 of the symptomatic individuals with mild malaria anaemia with OR 11.2 [95% (CI 2.2-56.9)] p=0.004 and OR 11.8 (95% CI 2.04-68.06) p=0.006 respectively after adjustment with age and sex while IL-12(p70) was, however, not statistically significant OR 1.28 [95% CI (0.32-5.13)] p=0.724. Ratios of pro- and anti-inflammatory cytokine responses Since the balance between pro- and anti-inflammatory cytokine responses is crucial during P. falciparum malaria infection, we determined the ratios of IFN-γ/IL-10, IFN-γ/IL-12(p70) and IL-12(p70)/IL-10 within the study population. To make the distribution more symmetrical, logarithmic transformation of the ratios were applied (Figure 4). The estimated ratio of IFN-γ/IL-10 symptomatic children (SC) of the ratio IFN-γ/IL-10 asymptomatic individuals (AI) was statistically higher in symptomatic children with mean difference of 2.42 [95% CI (1.52-3.84)] p=0.0003 (Figure 4a). The estimated ratio of IL-12(p70)/IL-10 was however higher in asymptomatic individuals with mean difference of 0.44 [95% CI (0.27-0.7)] p=0.0009 (Figure 4b). Also, the estimated ratio of IFN-γ/IL-12(p70) was significantly higher in the symptomatic children with mean difference of 5.52 [95% CI (2.8-10.89)] p<0.0001 (Figure 4c). Discussion In this study, the levels of IL-10 and IFN-γ were significantly higher in the symptomatic children with uncomplicated malaria unlike the asymptomatic individuals who had mostly Th1 type of responses, though the mean age of the later was higher. The association of significant levels of IFN-γ seen in symptomatic children less than 6 months with malaria suggests that IFN-γ may be instrumental in immune-protection against malaria by limiting parasite replication. Though none of the subjects in the present study had severe malaria, our results showed significant levels of IFN-γ responses in symptomatic children <6months. In uncomplicated malaria, IFN-γ might help to limit parasite progression and activated CD8 T cells produce IFN-γ through macrophage activation17 and appears very important in limiting the disease progression to severe malaria with life threatening complications18. Previous study had shown the protective effect of IFN-γ responses to P. falciparum malaria in children living in malaria endemic regions19. Also, the ratio of IFN-γ/IL-12 in this study was statistically higher in symptomatic children further emphasizing the role of inflammatory responses in the limitation of parasite replication. Moreover, the antagonistic regulation of Th1 and Th2 cytokines in relation to intracellular parasites may explain the high ratio of IFN-γ/IL-10 observed in these symptomatic children with the view of regulating the cytopathic effects20 Earlier studies have expressed a possible pivotal role of IL-10 in the modulation of pro-inflammatory cytokine response of peripheral blood mononuclear cells during malaria infection21. Elevated levels of IL-10 have been reported in severe malaria22?and have been associated with less effective clearance of P. falciparum parasites23?due to its down-regulatory activity of anti-parasite inhibitory immune function24 IL-10 inhibits the production of IL-12 from monocytes24,25 and low IL-12 activity has been associated with severe P. falciparum malaria11,26 Recently, data from Mali suggests that heterozygousity in IL12B is associated with reduced expression of the gene encoding IL12p40 and a low level of IL12p70 production in cerebral malaria27. Another study suggests that the genetic variation in IL12B does not explain differences in parasitaemia in individuals28. However, the low levels of IL-12(p70)/IL-10 ratio observed among the symptomatic children with uncomplicated malaria in this study may characterize the suppression of the protective effects of IL-12 in view of the marked differences seen with the asymptomatic individuals? Our data indicate significant levels of antibodies in individuals above 5 years of age than those under 5 years. However, high IgM mean concentration level was more associated with parasitaemia than age. Significant levels of IgG1 and IgG3 subclass antibodies were seen in individuals above 5 years while IgG3 and IgG4 were notably low in <5 years. IgG antibodies are acquired from the maternal circulation during gestation, lost five to seven months after birth and protect the young infant from severe malaria manifestations by slowing the rate of intraerythrocytic parasite multiplication29 Immunoglobulins; IgG1, IgG3 and IgG4 readily cross the placenta and also play important roles in protecting the developing foetus. IgG3 has a short life and is the most effective complement-activator followed by IgG1 and IgG2. The levels of IgG1 and IgG3 to the malaria antigen suggest an interdependent induction of these two subtypes but there was no clear pattern with malaria-specific IgG4. The systematic development of anti-malarial IgG1, IgG2, IgG3 and IgG4 in response to repetitive exposure of these individuals to P. falciparum malaria infection signifies an age-related development of their responses. IgG1 antibodies, being the most abundant subclass, seem to first initiate parasite density control in view of marked immunoglobulin geometric differences observed in this study suggesting that it plays significant role in opsonisation and possibly complement-mediated lysis of free merozoites and/or infected red blood cells. IgG1 and IgG3 antibodies, in humans, have high affinity for Fc receptors, interacting through complement fixing, phagocytosis, and IgG3 appears most competent in phagocytosis and mediate FcR cross-linking30. The low levels of anti-P. falciparum IgG3 antibodies observed in the symptomatic children (especially <5 years) showed an insufficient cumulative exposure to the malaria antigens, thus may affect the protective effects of this subclass in children living in malaria endemic region. In view of this, the anti-parasite immunity seen in asymptomatic individuals seems to reflect quantitative switch to specific IgG subclasses31. However, different malaria antigens have been shown to induce different IgG subclass profiles despite the fact that they are apparently present in the immune system under identical conditions32. Conclusion Malaria infection has a huge effect on the delicate balance between the inductions of pro- and anti-inflammatory cytokines, which in turn determines the nature of the protective or pathogenic effect subsequently induced. The differences in the ratio between Th1-and Th2-cytokine in adults in this study reflects their capability to control and limit parasite multiplication in view of marked differences observed within the children presenting with malaria. Though the levels of immunoglobulin responses differed with blood-stage P. falciparum antigens, the immunoglobulin subclasses gradually increased with advancement in age and varied with malarial antigens. Meanwhile, the low levels of malaria specific antibodies observed in the symptomatic children especially <5 years explicate their susceptibility to clinical episodes of malaria due to poor specific immune responses. Acknowledgements We acknowledge all the participants in this study and their parents/guardians. We thank the clinical staff members of Oni Memorial Hospital. We also appreciate with sincere gratitude the efforts of Drs Femi Ibrahim and K. Akpala in this project. N. C. Iriemenam was a recipient of ICSC World Laboratory and UNESCO-IUMS-SGM scholarships and C. M. F. Okafor received the MIMPAC network Exchange student award to Stockholm University. References
Copyright © 2009 - Makerere Medical School, Uganda The following images related to this document are available:Photo images[hs09014t1.jpg] [hs09014f4.jpg] [hs09014t2.jpg] [hs09014f3.jpg] [hs09014t3.jpg] [hs09014f1.jpg] [hs09014f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}