 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
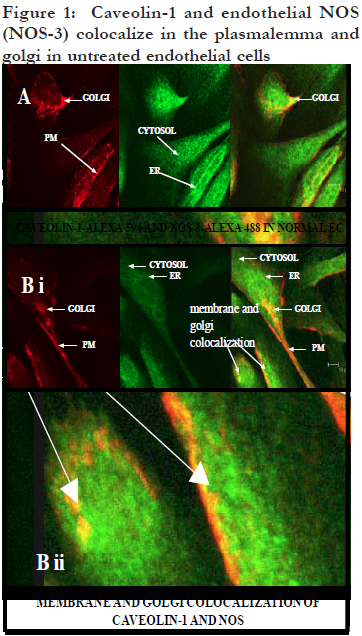
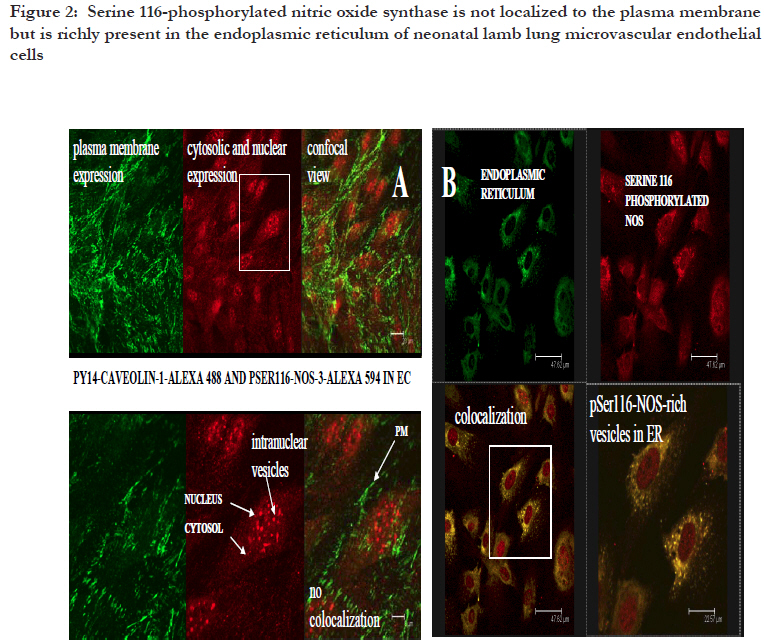
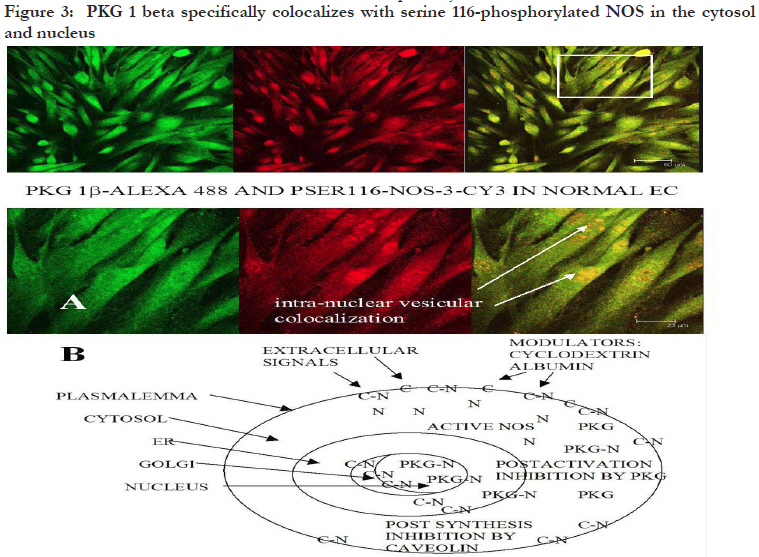
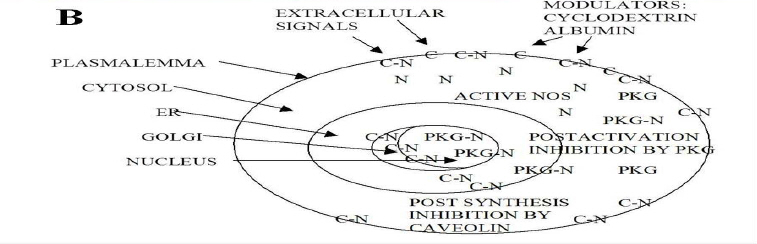
African Health Sciences, Vol. 11, No. 4, Dec, 2011, pp. 526-534 Inhibitors caveolin-1 and protein kinase G show differential subcellular colocalization with Nitric oxide synthase *Adebola TJ1 , Usha Raj2 1. Department of Pharmacology, Lagos State University College of Medicine. 1 Oba Akinjobi Way, GRA, Ikeja, Lagos, P.M.B. 21266, Ikeja, Lagos, Nigeria. Code Number: hs11108 Abstract Background: Nitric oxide synthase (NOS) is negatively regulated by protein-protein interactions with caveolin-1 before extracellular activating signals release it for nitric oxide (NO) production. Smooth muscle protein kinase G (PKG) is a down-stream effector of NO signaling for relaxation of vascular smooth muscle cells (SMC). The PKG is also found in endothelial cells and it inhibits activated NOS within endothelial cells. Key words: caveolin-1, protein kinase G, nitric oxide synthase Introduction Nitric oxide synthase (NOS) produces nitric oxide (NO) which is important for vasodilatation and other vascular functions.1,2,3 Low levels of NO have been associated with endothelial dysfunction and diseases such as pulmonary hypertension, atherosclerosis, heart failure, and sexual dysfunction.4,5 Many factors modulate NOS function6,7,8 but we still know very little about how the basal function of NOS is sustained. We recently reported that protein kinase G (PKG) inhibits NOS.9 In various reports, we speculated that PKG in endothelial cells is involved in the termination of NOS activity: its interaction with NOS involves intracellular vesicular systems and the nucleus;9 it inhibits both basal and stimulated nitric oxide synthesis;10 it appears to involve both phosphorylation of serine 116 and decreased phosphorylation of threonine 1177, as well as molecular change of NOS molecule;11 and it may lead to degradation of NOS and the recycling of a NOS fragment that may act as a transcription factor for de novo enzyme synthesis.12 A regulator of NOS that has been well studied is caveolin-1. From published work, immunoprecipitation of NOS from bovine lung microvascular endothelial cells resulted in coprecipitation of caveolin-1.13 Caveolin-1 is well known to be abundant in the membrane region14,15 and is synthesized and packaged16,17,18 or recycled19,20 in the ER/Golgi network. Caveolin-1 is the structural protein of caveolae which are invaginations of the plasma membrane, but it also found in the cytosol and other parts of the cell. The binding of caveolin1 to NOS has been shown to negatively regulate NOS activity.21,22 It has been suggested that caveolin1 may play the role of a molecular chaperone that directly inactivates NOS23, and some studies support the binding of NOS and caveolin-1 by in vitro/ex cyto approaches. However, colocalization of caveolin-1 and NOS in the cytosol has not been demonstrated, therefore caveolin-1 may not have a role as a chaperone in NOS metabolism. In the present study, we investigated indirectly whether under basal conditions, PKG is a differential inhibitor of NOS compared to caveolin1, a better known negative regulator of NOS.24 The research question was: do PKG and Caveolin-1 colocalize with NOS differentially? This can indicate that their known roles as inhibitors of NOS differ. To do this, the subcellular distribution and colocalization of NOS with that of caveolin-1 and PKG were compared. Therapeutic modulation of NO production has been attempted by diverse pharmacological approaches.25 Understanding endogenous regulation of basal NOS activity and the metabolic cycle of the NOS enzyme can help us develop precise therapeutic strategies for regulating basal release of NO in disease conditions where NO is depleted, or when there is excessive production of NO. Our hypothesis is that caveolin-1 and PKG regulate NOS at different stages of NOS life cycle. Based on results from previous studies,9,10,11,12 we speculate that while caveolin-1 renders nascent NOS inactive pending activation signals, PKG deactivates active NOS and subjects the spent enzyme to degradation. If this were true, the two inhibitors would colocalize with NOS at their respective sites of inhibition. In this study, the subcellular distribution of caveolin-1 and PKG and their colocalization with NOS in ovine lung microvascular endothelial cells are compared using fluorescence immunohistochemistry and confocal microscopy. Methods Animals This work was reviewed and approved by the Animal Care and Use Review Committee of Los Angeles Biomedical Research Institute. Newborn lambs aged <2 were obtained from Nebeker Ranch (Lancaster, CA). Lambs were anesthetized with 30 mg/kg ketamine HCl (Phoenix, MO) and then sacrificed with pentobarbital (Virbac, TX) overdose. Lungs were dissected out and transported in pre-gassed (100% O2) ice cold Krebs’ solution (salts were from Sigma, MO). Isolation of ovine lung microvascular endothelial cells (LMVECs). Cells were isolated and characterized as previously reported.26 Miniature explants of extreme lung edge parenchyma were gently crushed with a hemostat to release endothelial cells into cell culture medium (DMEM (Invitrogen, NY) plus 10% FBS (Atlanta Biologicals, GA), 1% Gibco® antibiotic/antimycotic mixture (Invitrogen)) under sterile conditions. Explants were separated with Steriflip® vacuum filtration units and the cells were distributed in culture wells and incubated under normal conditions (in a HEPA filtered incubator, Thermo Electron, Ohio, at 37 °C with 5% CO2 mixed in air). Derived cultures were characterized by immunohistochemistry to show expression of CD31 and von Willebrand Factor as previously reported26. In the course of the experiments, the primary cells were passaged up to five times. Detection of subcellular colocalization of NOS with its negative regulators caveolin-1 and PKG by immunohistochemistry Quiescent investigation (no treatment or intervention) of microvascular endothelial cells were done. Routine immunohistochemistry as previously reported9 was performed on early passage (2-3) cells grown on Lab-Tek® II 4-chamber slides (Nalge Nunc, Naperville, IL). In each set of chambers, one chamber was used as the primary antibody blank to test for non-specific binding and extraneous fluorescence and three were used as triplicates of the same investigation with the primary antibody. Before fixation with 4% paraformaldehyde, cells were kept as quiescent as possible to minimize shear stress. Cells were permeabilized with 0.2% triton X-100 and blocked with 20% bovine serum albumin. Highly specific monoclonal or affinity purified rabbit polyclonal or goat primary antibodies were used. Colocalization of NOS and caveolin-1 Primary antibodies in dilutions of 1: 200 were used against caveolin-1 (rabbit, Abcam, Cambridge, MA), NOS (mouse, Abcam, Cambridge, MA), tyrosine 14 phosphorylated caveolin-1 (mouse, BD Translab, San Jose, CA), serine 116 phosphorylated NOS (rabbit, Sigma, St Louis, MO), and endoplasmic reticulum marker protein disulfide isomerase (PDI) (mouse, Invitrogen, NY). Secondary antibodies were used in dilutions of 1: 125. To investigate NOS and caveolin-1 colocalization or pSer116NOS and pY14caveolin colocalization, cells were counterstained with F(ab’)2 fragment specific secondary antibodies (Invitrogen, NY) - one raised in goat against rabbit and conjugated to alexa 594 and the other raised in goat against mouse and conjugated with alexa 488. For pSer116NOS and PDI colocalization, cells were counterstained with F(ab’)2 fragment specific secondary antibodies (Sigma, St. Louis MO) raised in sheep, one against rabbit and conjugated with cy3 and the other against mouse and conjugated with FITC. Colocalization of NOS and PKG For pSer116NOS and PKG 1beta colocalization, primary antibodies in dilutions of 1: 200 were used against serine 116 phosphorylated NOS (rabbit, Sigma, St Louis, MO) and PKG 1beta (goat, Santa Cruz, CA). Cells were counterstained with a donkey anti-goat alexa 488 conjugated antibody (Invitrogen, NY) plus a F(ab’)2 fragment specific sheep anti rabbit cy3 conjugated secondary antibody. Microscopy Mounted slides were studied by fluorescence microscopy using a Zeiss Axioscop 40 microscope, and epitope colocalization was recorded by confocal microscopy using a DMIRE2 confocal inverted microscope and Leica v2.61 confocal software. Water immersion objective lenses (x63) were used to take confocal images. Specific lasers, scanning parameters and specific excitation wavelengths were set to match fluorochromes observed. For setting of filters, the emission range for green fluorescence tags (alexa 488 and fluorescein (FITC)) was 500560 nm, and for red fluorescence tags (alexa 594 and cy3) the range was approximately 600-700 nm. Results Differential sites of colocalization of caveolin-1 or PKG with nitric oxide synthase in untreated ovine lung microvascular endothelial cells Caveolin-1 marked with red alexa 594 was clearly concentrated in the plasma membrane and Golgi regions of the lung microvascular endothelial cells, but was also in the cytosol [Figure - 1]. The NOS, marked with green alexa 488, was ubiquitous all over the cytosolic and nuclear domains of the untreated cells with greater intensity in the ER network [Figure - 1]. Caveolin-1 and NOS were colocalized in the Golgi region and regions of the plasma membrane [Figure - 1] and not in the cytosol. Serine 116 phosphorylated NOS marker was found in the cytosol and distinctively in nuclear region vesicles marked with red alexa 594 [Figure - 2A] and endoplasmic reticulum vesicles marked with red cy3 [Figure - 2B]. The PKG1beta was distributed all over the cytosolic and nuclear domains of the untreated cells without a greater intensity in any region [Figure -3A]. PKG 1beta was intensely colocalized with serine 116 phosphorylated NOS in the nuclear domain in vesicular structures [Figure - 3A]. Such colocalization was also apparent in diffuse expression in the cytosolic domain of the cells [Figure - 3A] [Figure - 1]. Triple monolayers of passage 2 neonatal ovine lung microvascular endothelial cells were fixed with 4% paraformaldehyde, permeabilized with 0.2% triton X-100, blocked with 20% bovine serum albumin, and then incubated with affinity purified rabbit anti caveolin-1 (1: 200) and monoclonal anti NOS(1: 200) primary antibodies overnight at 4C. They were counterstained with 1: 125 dilutions of F(ab’)2 fragment specific goat secondary antibodies against rabbit conjugated to red alexa 594, and against mouse conjugated to green alexa 488. Pictures were taken with a DMIRE2 confocal inverted microscope and Leica v2.61 confocal software. Arrows show areas of caveolin-1 and NOS co-localization seen in random fields from two monolayers (A and B i) with enlargement of some membrane portion of B to show numerous minute membranous colocalization foci (yellow, B ii) (scale in B i shows10 micro meters). [Figure - 2] Part A shows that plasmalemma associated tyrosine14-phosphorylated caveolin-1 and serine 116-phosphorylated NOS are differentially located in endothelial cells. Triple monolayers of untreated passage 2 neonatal ovine lung microvascular endothelial cells were fixed with 4% paraformaldehyde, permeabilized with 0.2% triton X-100, blocked with 20% bovine serum albumin and then incubated with mouse anti tyrosine phosphorylated caveolin-1 (1: 200) and rabbit anti serine116-phosphorylated NOS (1: 200) primary antibodies for 2h at room temperature. They were counterstained with 1: 125 dilutions of F(ab’)2 fragment specific goat secondary antibodies against mouse conjugated to green alexa 488, and against rabbit conjugated to red alexa 594. Pictures of a field representative of all three monolayers are shown taken with a DMIRE2 confocal inverted microscope and Leica v2.61 confocal software. Left panels show separation of pY14-caveolin-1 marker distinctively in plasma membrane and pSer116-NOS marker distinctively in the cytosol and nuclear region. Insert from upper left panel shown in lower left panel depicts the distribution of the pSer116-NOS moiety in nuclear region vesicles. Part B shows that serine 116-phosphorylated NOS is found in vesicular structures in endothelial cell ER and nuclear regions. Triple monolayers of untreated passage 3 neonatal ovine lung microvascular endothelial cells were fixed with 4% paraformaldehyde, permeabilized with 0.2% triton X-100, blocked with 20% bovine serum albumin and then incubated with mouse anti protein disulfide isomerase (1: 200) (an endoplasmic reticulum (ER)-specific marker) and rabbit anti serine116-phosphorylated NOS (1: 200) primary antibodies for 2h at room temperature. They were counterstained with 1: 125 dilutions of F(ab)2 fragment specific sheep anti mouse conjugated to green FITC and sheep anti rabbit conjugated to red cy-3 secondary antibodies. Pictures were taken with 116 phosphorylated NOS moiety passes through the a DMIRE2 confocal inverted microscope and Leica ER and nuclear regions of quiescent cells in certain v2.61 confocal software. Insert shows that a serine processing vesicles indicating a NOS metabolic pathway. [Figure - 3] Part A shows that PKG1beta and serine-116phosphorylated NOS moiety colocalize in the cytosol and nuclear region vesicles. Quadruplet monolayers of untreated passage 3 neonatal ovine lung microvascular endothelial cells were fixed with 4% paraformaldehyde, permeabilized with 0.2 % triton X-100, blocked with 20% bovine serum albumin and then incubated with 1: 200 dilutions of goat anti PKG 1beta and rabbit anti serine116phosphorylated NOS primary antibodies for 2h at room temperature. They were counterstained with 1: 125 dilutions of alexa 488-conjugated donkey anti goat and cy-3 conjugated F(ab)2 fragment specific sheep anti-rabbit secondary antibodies. Pictures were taken with a DMIRE2 confocal inverted microscope and Leica v2.61 confocal software. The PKG 1 and pSer116-NOS markers colocalized in the cytosol and intensely in vesicles in the nuclear region. Part B is a hypothetical schema of differential subcellular co-localization of caveolin-1 or PKG1beta as inhibitors of NOS in endothelial cells. Caveolin-1 co-localizes with and is known to inhibit nascent NOS produced in the Golgi or supplied to the plasma membrane for extracellular signals. The figure depicts caveolin-1 bound to NOS in the Golgi network and plasmalemma as [Figure - 1] indicates and as previous workers have shown.13,21,22 The figure also depicts that PKG inhibits NOS after it is activated in the cytosol and is involved in the recycling of spent NOS, as [Figure - 3] indicates. By some mechanism, PKG colocalizes with NOS or with a NOS fragment in the nuclear region. Key: N = free nitric oxide synthase; C = free caveolin-1; C-N = hypothetical caveolin-1-NOS complex; PKG-N = hypothetical protein kinase G-NOS complex. Discussion In the present study, immunofluorescence staining and confocal microscopy were used to determine colocalization of molecules of interest within endothelial cells. “Colocalization …occurs when fluorescently labeled molecules bind to targets that lie in very close or identical spatial positions. The application of highly specific modern synthetic fluorophores and classical immunofluorescence techniques, coupled with the precision optical sections and digital image processing horsepower afforded by confocal and multiphoton microscopy, has dramatically improved the ability to detect colocalization in biological specimens.”27 We were able to establish that caveolin-1 and protein kinase G respectively colocalized with nitric oxide synthase in quiescent endothelial cells but at different subcellular sites. The pictures shown here are randomly selected representative fields. In [Figure -1] the immunostaining picks up cellular regions of intense appearance of either caveolin-1 or NOS individually. It also depicts regions where their colocalization was intense within the normal cellular architecture. The distributions were observed in cellular regions/domains - plasma membrane regions, golgi regions -as stated in the results, but multiple staining was not used to make the observation in these initial studies which were kept as quiescent and untreated as possible. Caveolin-1 was distributed in the cytosol and was intensely expressed in the plasma membrane and golgi regions of the cell [Figure - 1]. Definite colocalization of caveolin-1 and non-phosphorylated NOS was observed at the plasma membrane and Golgi regions of the cells by double staining [Figure - 1) and this goes well with the known roles of caveolin-1 as a negative regulator of nascent NOS before it is activated by extracellular signals.21,24 As discussed in the background, colocalization of caveolin-1 and NOS in the cytosol has not been demonstrated and therefore caveolin-1 may not have a role as a chaperone in NOS metabolism. We also, in this present work, were unable to co-localize caveolin-1 with NOS in the cytosol. On the other hand, we colocalized PKG with NOS in the cytosol. Compared to caveolin-1, PKG as an inhibitor of NOS under physiological conditions has not been extensively studied. In a recent publication, we showed that the serine 116 site of NOS plays a role in termination of NOS activity and that protein kinase G acts as a mediator of NOS inactivation.9 In this current study, we show that the subcellular colocalization of PKG and NOS indicate that PKG interacts with NOS that is not in the plasmalemma. The plasma membrane is the entry point of extracellular signals that can activate NOS. The nucleus is well known to be the site of transcription in the cell and the ER and Golgi network are considered the sites of degradation/synthesis and processing of various types of molecules and biological packages. The ER extends generally from the nuclear boundary but may traverse the cytosol in certain cell types as it appears in these neonatal ovine LMVECs. Whereas NOS can be seen distributed uniformly all over the cell with some intensity in the ER-Golgi network [Figure - 1], serine 116 phosphorylated NOS shows distinct intense vesicular distribution in the ER-nuclear region apart from its punctuate cytosolic appearance [Figures - 2] and [Figure - 3]. Interestingly, it is this serine116-NOS that PKG colocalizes with [Figure - 3] in the different subcellular compartments, suggesting a chaperone complex with the two molecules. There are numerous integral molecular components of the plasma membrane. Since caveolin-1 is an integral membrane protein of caveolae14,15 found all over the membrane and tyrosine14 phosphorylated caveolin-1 is associated with focal adhesion points28,29 which are also membrane areas, in this study the antibody against tyrosine-14 phosphorylated caveolin-1 was used to show membrane regions of the cellular monolayers. Caveolin-1 negative regulation of NOS can be released by activating stimuli such as calcium/ calmodulin or bradykinin30,31 making active NOS available in the cytosol. Stained against plasmalemmaassociated,tyrosine-14-phosphorylated caveolin-1, pSer116NOS is clearly not localized to the plasmalemma [Figure - 2A] as expected of the preactivation nascent NOS. This may be contrasted with [Figure - 1] where NOS colocalized with caveolin1 in the plasma membrane. On the other hand, the present work also clearly shows that phosphorylated serine 116 region of NOS is detected in the cytosolic region, and that there is a distinctive appearance of this marker in vesicles both in the endoplasmic reticulum [Figure - 2B] and in the nuclear region [Figure - 2] and [Figure - 3]. This may indicate that pSer116NOS is a post activation form, possibly a spent form, of NOS. Immuno-presence of serine 116 phosphorylated portion of NOS in the nuclear region is seen in [Figure - 2] but we cannot tell if it represents the whole enzyme molecule or an Nterminal degradation product. This nuclear region presence is probably physiological and it may indicate a life cycle of the enzyme involving serine 116 phosphorylation and the nucleus. We cannot say whether NOS localizes to the nucleus in a phosphorylation-dependent manner. We need to investigate whether a serine 116 phosphorylated translocated NOS or a serine 116 phosphorylated NOS N-terminal fragment is involved in de novo transcription under basal NO production activity. Recently we published some fluorescence activated cell sorter analyses of the interaction of PKG and activated NOS, showing that PKG causes deletion of the carboxy terminal of NOS, possibly a degradation process.11 Furthermore, we showed that in western blot analysis of whole cell lysates, serine 116 phosphorylated NOS appears in molecular weight fragments of about 60, 50 and 35 kDa, and that the N-terminal remnants of NOS may be normally found in control endothelial cells and may be increased by PKG activation.12 From the present data, it appears that serine 116phosphorylated NOS or NOS fragment appears to enter a recycling process since the in-cell immunostaining pattern is all diffuse, punctuate, and vesicular in appearance. We need to perform further molecular studies to trace the apparent translocation of the suspect NOS fragment from the cytosol into the ER network and finally into the nucleus as can be inferred from [Figure -2B] and [Figure - 3A]. This would be a novel hypothesis and biologic pattern and we are not yet able to comment on it mechanistically but present here the interesting pictorial findings. Moreover, we would need to re-establish the finding in available human cells. Comparing the two physiological inhibitors of NOS, caveolion-1, and PKG, PKG may be involved in a pro-translocation phosphorylation of NOS for degradation of the enzyme and a carboxyterminal deleted fragment of NOS, possibly on a PKG-1â-complexed chaperone, may be processed and recycled via the ER-nucleus network. Caveolin1 has not been demonstrated to colocalize naturally in the cytosol with NOS and cannot be attributed a chaperone function. A major question that can now be raised with respect to NOS metabolism is whether a degradative process for spent NOS that is linked with de novo transcription and synthesis of NOS to form a selfsustaining cycle for continual NO generation in endothelial cells exists. We do not know how NOS is continually replenished to sustain basal NO production under physiologic conditions, a vital function in the cardiovascular system. Conclusion Caveolin-1 and PKG, which are known to inhibit NOS activity, colocalize with NOS at different subcellular sites. They may therefore inhibit NOS at different phases of the metabolic cycle of NOS. It appears that caveolin-1 exerts a post-translational inhibition on nascent NOS at the plasma membrane and within the Golgi and makes NOS available for extracellular signals [Figure - 5].32,33 Differently, PKG appears to exert a post-activation inhibition on NOS and is associated with NOS (possibly NOS Nterminal fragments) in the cytosolic domain and within ER and nuclear-site processing vesicles [Figure - 3]. The role of PKG in endothelial NOS deactivation10 and recycling needs further studies against the background of the caveolin-1-NOS calcium-dependent association/dissociation24,34,35 events being considered as the main mechanism for negative regulation of endothelial NOS. Acknowledgment This work was done at the Division of Neonatology, Department of Pediatrics, Los Angeles Biomedical Research Institute at Harbor UCLA Medical Center (LABiomed), Torrance, California, USA, and was supported by United States of America Government grant MIRS HL059435 to T. A. John under the supervision of Prof. J. U. Raj. The authors are very grateful for the expertise of Jennifer Dedes, core operator of the confocal microscopy and other facilities at the LABiomed for her invaluable contribution to generating images for this paper. References
Copyright © 2011 - African Health Sciences The following images related to this document are available:Photo images[hs11108f1.jpg] [hs11108f3.jpg] [hs11108f2.jpg] [hs11108f5.jpg] [hs11108f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}