 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
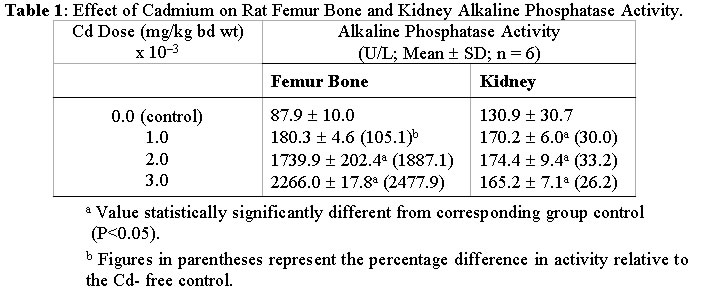
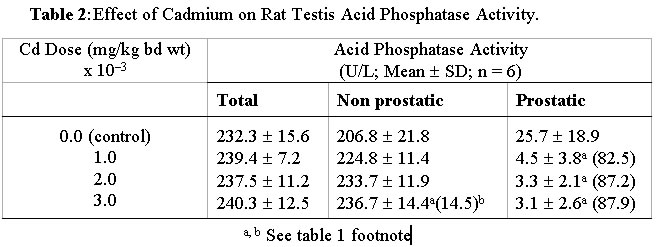
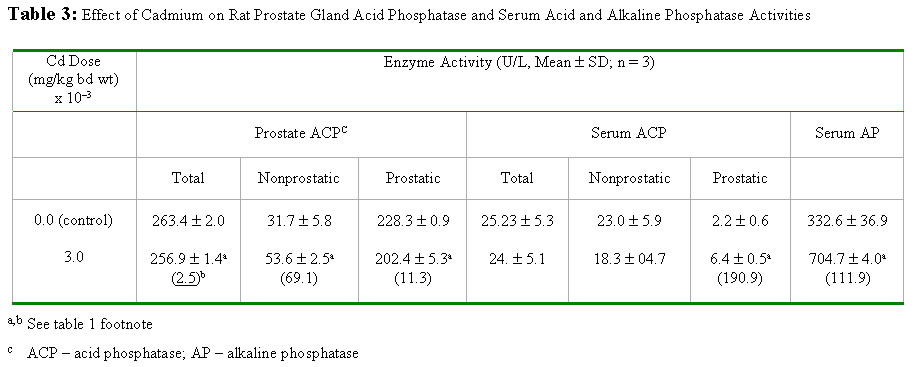
Journal of Applied Sciences & Environmental Management, Vol. 6, No. 1, June, 2002, pp. 9-13 Effects of Cadmium Exposure on Bone and Kidney Alkaline Phosphatase Activity and Acid Phosphatase Activity in Testis and Prostate Gland in the Male Rat *OBI, F. O.; ILORI O. OLABODE Department
of Biochemistry Faculty of Science University of Benin, Benin City, Edo State
NigeriaEmail: obifo@uniben.edu.ng Code Number: ja02002 ABSTRACT: This paper examines the effects of varying doses of cadmium on bone and kidney alkaline phosphatase and on testis and prostate acid phosphatase after 4 weeks of administration to separate groups of rats. Relative to the cadmium-free control rats femur bone alkaline phosphatase activity was significantly (P<0.05) elevated in rats exposed to 2 mg cadmium /kg body weight and in those exposed to 3 mg/kg. Compared to control rats kidney alkaline phosphatase activity was also statistically significantly (P<0.05) elevated in separate groups of rats exposed to 1, 2 and 3 mg cadmium/kg body weight. Prostatic acid phosphatase activities in the testis and prostate were significantly (P<0.05) decreased in the groups of rats separately exposed to 1, 2 and 3 mg cadmium/kg. At the highest dose nonprostatic acid phosphatase activity increased significantly (P<0.5) in testis and prostrate. In serum at the highest dose nonprostatic acid phosphatase decreased while the prostatic isoform and alkaline phosphatase increased significantly (P<0.05). Our results indicate that cadmium enhances the activities of alkaline phosphatase in the bone and kidney, which of nonprostatic acid phosphatase in the testis and prostate but lowers the activity of the prostatic isoform in both.@ JASEM Cadmium (Cd), a heavy metal is toxic to both humans and animals. It is widely distributed in the general environment of today as a result of its diverse use in industries (Nicholson et al, 1983; Friberg et al, 1986). A widely reported case of chronic Cd intoxication in humans is the itai-itai disease which was reported as endemic to a heavily Cd-polluted area of Toyama Prefecture, Japan (Kasuya et al, 1992; Yasuda et al, 1995). The pathogenetic mechanisms of the disease is yet to be completely elucidated. However, the general understanding of the course of the disease is that the cadmium accumulates in the renal tubular cells and leads to gradual impairment of function because of its toxic effect. Subsequently severe osteomalacia ensues due to loss of minerals such as calcium, phosphates and bicarbonates into the urine (Horiguchi et al, 1994). Experiments involving calcium deficient and cadmium supplemented diet and water have been done using animal models in order to induce osteomalacia (Itokawa et al, 1974; Takashima et al, 1980). Such studies on osteomalacic changes in femur induced by long-term administration of Cd supplemented diet to rats revealed the existence of osteoid tissues containing uncalcified glycoaminoglycans and collagenous tissues (Takashima et al, 1980). Besides the kidneys and bones another organ that is susceptible to the toxic effect of cadmium is the testis (Gabbiani et al, 1974; Francavilla et al, 1981; Jones et al, 1988; Draper and Timbrel, 1996). In male mice cadmium causes marked necrosis of seminiferous tubules as well as interstitial hemorrhage and edema (Jones et al, 1988). In rats there is massive loss of creatine from the testis leading to significant creatinuria (Draper and Timbrel, 1996). The mechanism by which cadmium triggers demineralisation of bones leading to osteomalacia and damages other organs and tissues is yet to be adequately clarified. We are inclined to believe that further understanding of the changes induced by cadmium which lead up to osteomalacia and other reported effects on target organs may emerge by examining its effect on enzymes endogenous to the target organs. This has not received satisfactory attention and so we have focused our studies on alkaline phosphatase (EC 3.1.3.1) an enzyme found in the bone and kidney and a related enzyme, acid phosphatase (EC 3.1.3.2) found in the testis and prostate gland. Alkaline phosphatase is richly produced by osteoblasts in the bone; and is suspected to be involved in the transport of phosphate across cell membranes and calcification of bone matrix (Verley, 1967; Nakamura et al, 1988). MATERIALS AND METHODS Animals: Thirty post weaning healthy male rats (Rattus nervegious; Wistar strain), weighing between 160-175g obtained from Lagos University Teaching Hospital Animal Unit were used for this study. They were given chow (Guinea Feed, Nig. Plc) and water ad libitum and left to acclimatise for a period of two weeks before the experiment was commenced. Chemicals and Reagents: Cadmium sulphate (3CdSO4.8H2O), chloroform, sodium chloride (E. Merck, Darmstadt, W. Germany). Acid and alkaline phosphatase assay kits (Randox Lab. Ltd. N. Ireland). Preparation of cadium sulphate solution: Cadmium sulphate octahydrate (3.76mg) was dissolved in distilled water and made up to one litre to give 1.20mg/l cadmium stock solution. From this stock appropriate dilutions were made with distilled water to obtain 0.80 and 0.40-mg Cd/l solutions of cadmium. Treatment: Experiment I: Experiment II: Animal sacrifice and Tissue Preparation: At the end of the 4th week, the rats were anaesthesized in a chloroform-saturated chamber (Expt. I and II) and exsanguinated by cardiac puncture (Expt. II). The hind limbs were disarticulated and the femur bones were removed, freed of traces of muscle tissue, weighed and stored at -20° C until required (Expt. I). The kidneys, testes (Expt. I) and prostate gland (Expt. II) were excised and stored at -20° C until required. The blood samples were left standing on ice to clot and the sera obtained by centrifugation, 5000 rpm for 10 min (Gallenkamp Bench Centrifuge). The sera were also stored at -20° C until required. The femur bones, the kidneys, the testes and prostate were homogenized in saline (1g/4ml) in an ice-cold motar. The homogenates were transferred into ice-cold centrifuge tubes and centrifuged at 5000 rpm for 10 min. The supernatants were separated and left at -20° C until required. Enzyme Assay: The activities of acid phosphatase in prostate gland, serum and testes were evaluated using the colorimetric method described by Bodansky (1972) while the method described by Plummer (1978) was used for alkaline phosphatase assay. The procedures outlined in the respective assay kit manuals (Randox Lab Ltd.) were adopted for the semimicroassay (alkaline phosphatase), for total and prostatic acid phosphatase determinations and for calculating the activities of the enzymes. To calculate alkaline phosphatase activity in units/litre, the formula used is 2760 x DA405 nm/min while the formulae for total and prostatic acid phosphatase are 101 x A405 sample 1 and 101 x (A405 sample 1 - A405 sample 2) respectively. Statistical Analysis: The data are presented as means mean value of the various treatment groups in experiment I were compared using ANOVA and the least square difference test (Lapin, 1978). The mean values in experiment II were compared using Student's t-test, two tailed analysis (Elzey, 1971). The significant level was set at P<0.05. RESULTS AND DISCUSSION Changes in femur bone and kidney alkaline phosphatase activities induced by varying doses of cadmium after 4 weeks are presented in Table 1. Table 2 shows the changes in testis total, nonprostatic and prostatic acid phosphatase activities due to exposure to the same doses of cadmium for the same period of time. The effects of cadmium at 3mg/kg body weight only on prostate gland and serum total, non prostatic and prostatic acid phosphatases as well as on serum alkaline phosphatase are shown in Table 3 In this short term study of chronic oral administration of cadmium to male rat a statistically significant (P<0.05) increase in femur bone and kidney alkaline phosphatase (AP) activities was observed particularly in rats exposed to 2 and 3 mg Cd/kg body weight. A corresponding increase in serum AP activity was also observed at the highest dose (Table 1). Prostatic acid phosphatase activities (ACP) in the testes decreased progressively as the dose of Cd increased. The nonprostatic ACP increased although not significantly above the control except at the highest dose (Table 2). Rich sources of AP are the bile canaliculli of the liver, osteoblasts in the bone, proximal tubules in the kidney and mucosal cells of the small intestine (Verley, 1967). Damage to any of these organs or tissues would lead to elevated activity of its isoform of AP in the serum (Ngaha et al, 1989; Lin and Wang, 1986). This enzyme along with others and a number of nonenzyme proteins such as N-acetyl-b-D-glucosaminidase (NAG) and b-2-microglobulin (b 2-MG) are currently being used as markers for Cd-induced renal damage (Kido et al, 1995). Of particular interest is urinary intestinal AP (IAP), which leaks from damaged mucosal cells and out into the urine via damaged kidneys. Moderate exposure to Cd is known to cause the leakage of IAP (Kido et al, 1995). The significantly high AP activity detected in the serum in this study could have come from any of the tissues/organs enumerated above, particularly the kidney and bone, which are established target organs of cadmium toxicity (Kido et al, 1995; Ahn and Park, 1995). However, there was no corresponding decrease in bone and kidney AP relative to the Cd-free controls, which would have been the case if they were merely damaged by this toxicant. It seems, therefore, that Cd induces the biosynthesis of AP in the bone and kidney before the disruption of cellular integrity, if any, and this usually occurs by way of lipid perodixation (Sarkar et al, 1995; Stacey and Klaassen, 1981). Relative to the bone Cd-induced increase in the activity of AP in the kidney is considerably lower and it was almost completely stimulated maximally in the latter organ by the lowest doses (Table 1). There are two possible interpretations of these observations. The first is that bone responds more readily to Cd-induced AP biosynthesis and/or enhanced activity than the kidney. Second interpretation is that the kidney does not respond to Cd-induced AP synthesis, rather it responds to changes brought about by Cd which enhance the activity of kidney constitutive AP. The latter appears to be a more plausible interpretation in view of the barely 26 - 33% increase in AP activity in the kidney relative to Cd-free control and the profound difference in bone AP activities between the Cd-exposed and Cd-free control that ranges from 105 - 2478% (Table 1). All the same, it is conceivable that the difference is a reflection of a possible leakage of the kidney isoform inspite of initial biosynthesis, if the integrity of the organ has been substantially compromised because of Cd-exposure. This cannot be ignored at the moment in view of the fact that other investigators have detected intestinal AP in the urine (Kido et al, 1995) even in humans moderately exposed to cadmium. The validity of the above interpretations would have to be confirmed by further investigations and besides, it would be interesting to find out whether increased AP activity in the two organs is the underlying basis of bone decalcification (Takashima et al, 1980) and decreased glycoaminoglycan (Itokawa et al, 1974; Savolainen, 1995) in osteomalacic condition. The activity of acid phosphatase (ACP) which is known to be rich in the testis and in one of its accessory glands, the prostate, was examined. In the prostate prostatic and nonprostatic ACP (Table 3) exhibited the same pattern of response in terms of increase and decrease only, as in the testis. Cadmium causes testicular damage (Jones et al, 1988; Draper and Timbrel, 1996). An acute dose of 2-4 mg Cd/kg can cause testicular necrosis in experimental animals. The necrosis is usually preceded by edema, decreased blood flow and ischaemia (Francavilla et al, 1981). In these studies testicular enzyme levels were not examined and the effect of Cd on the prostate gland was rarely mentioned. The alterations in the activities of nonprostatic and prostatic ACP in the testis and prostate in the manners indicated above may therefore, be an integral part of the mechanism(s) involved in Cd-induced testicular cell injury and necrosis. In serum prostatic ACP activity increased significantly (P<0.05) (Table 3) and in the prostate it decreased although not significantly. Evidently there has been leakage of prostatic ACP into the blood. This is indicative of Cd-induced damage to the prostate gland. REFERENCES
Copyright 2002 - Journal of Applied Sciences & Environmental Management The following images related to this document are available:Photo images[ja02002t2.jpg] [ja02002t1.jpg] [ja02002t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}