 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Applied Sciences & Environmental Management, Vol. 6, No. 2, Dec, 2002, pp. 49-58 Culture Characteristics of Candida sp in Waste Conversion: Implications for Single-Cell-Protein-Enriched Feed Supplement Production ADOKI, A Institute of Pollution Studies, Rivers State University of Science & Technology, P. M. B. 5080 Port Harcourt Code Number: ja02028 ABSTRACT: A strain of Candida sp. was isolated from ripe banana pulp during the preliminary phase of a process for the production of a protein-enriched feed supplement. Morphological and biochemical tests demonstrated that the strain, which was bipolar and elongated, was not capable of growth at 37ºC but grew at room temperature only on dextrose and fructose and was able to supply its amino acid requirements in culture. The efficiency of conversion of readily utilizable carbon sources in fruit wastes used for feed supplement production by the test strain was also studied. Results obtained showed that reducing sugars were most readily utilized compared to other soluble carbohydrates. Conversion efficiencies were high and averaged about 75% in unsupplemented substrates. Supplementation with organic carbon and inorganic nitrogen compounds resulted in greatly increased biomass yields. @JASEM Recently, there has been considerable emphasis on the world food crisis and on the availability of 'waste' products, which could be utilized for alleviating food shortages (Bhattacharjee, 1970). Many of the developing countries where major nutritional problems exist produce an excess of materials rich in carbohydrates that can be utilized in fermentation processes to produce microbial protein which in turn can be used to upgrade both human and animal feeds. Traditional protein sources are relatively more expensive and the absence of well-developed technological facilities has contributed to losses incurred through spoilage of even the limited available sources. Consequently, there is the need to explore alternative ways of meeting the protein demands. Compared to the developing countries (mainly the U.S.A., U.K.& Japan) the less developed countries appear to have more abundant supply of agro-waste substrates that could be converted to additional protein sources (Moo-Young, 1977). Therefore, it is the aim of this study to demonstrate the utilization of fruit wastes for production of microbial protein that would serve to supplement the available traditional protein sources. Based on this, an attempt was made to produce single-cell-protein- enriched (SCP) feed supplement from a Candida sp. Using wastes derived from orange (Citrus sinensis), banana (Musa paradisiaca) and plantain (Musa sapientum). The choice of these raw material sources was based on the fact that wastes derivable from them, for example, as obtains in the processing of orange for juice or jam production are enormous and frequently create environmental problems associated with non-effective disposal systems. Similarly, a lot of wastes from these fruits are produced as a result of spoilage of harvested fruits. This is due to the dearth of adequate storage facilities. The magnitude of such spoilage could be well appreciated when local markets are surveyed. The microbe, Candida sp. was chosen because of its relatively very good isolation and growth on carbohydrate containing media tested. Similarly, its energy requirements were considered to be minimal as it grew very well at room temperature. MATERIALS AND METHODS Isolation: The yeast strain used for this study was isolated from ripe banana. Fifty grams of fresh ripe banana pulp was marshed in a pre-sterilized mortar (sterilized by autoclaving at 121ºC) and transferred aseptically to 100ml sterile distilled water in a 250-ml conical flask stoppered loosely with a cotton wool plug. The marshed banana mixture was left to stand for 48h and a loopful cultured on streptomycin-containing potato dextrose agar (PDA) as described by Cruickshank et al. (1975). The petri dishes were incubated at 25ºC for 24h. Pure culture of the desired strain was obtained after several streaks on the antibiotic containing PDA. Stock cultures of the isolated strain were maintained on agar slants of streptomycin-containing PDA in screw-capped bottles and stored at 4ºC. Morphological and Biochemical Characterization of Test Strain: Morphology: The morphological characteristics of the isolate were determined by noting colony size, shape, colour and cellular morphology by microscopy. Biochemical characterization: As an aid to further identification of the production strain some biochemical tests were carried out as outlined below. a.
Utilization of carbon and energy sources: Biochemical tests involving the utilization of dextrose, fructose, maltose, mannitol, lactose and sucrose were carried out as described by Cruickshank et al. (1975) and Bradshaw (1979). Inoculation of flasks: Flasks when cool were inoculated with 1ml portions of a suspension of yeast cells in sterile distilled water. These were then incubated in an orbital shaker set at 150 rpm at room temperature (25 - 28ºC). Determination of Sugar Utilization (Conversion) Efficiency
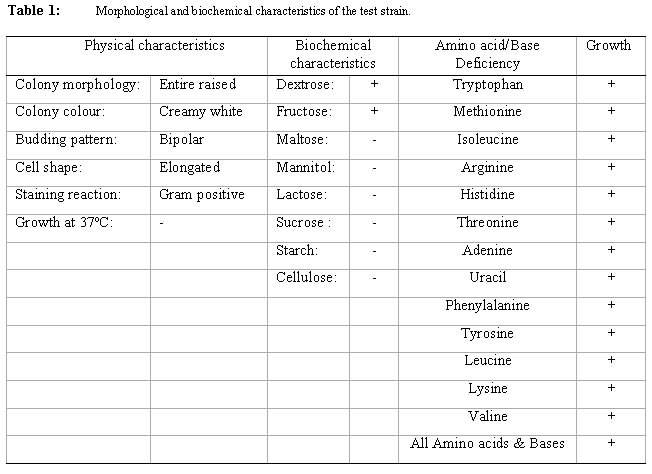
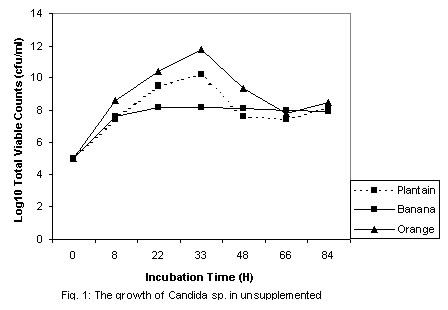
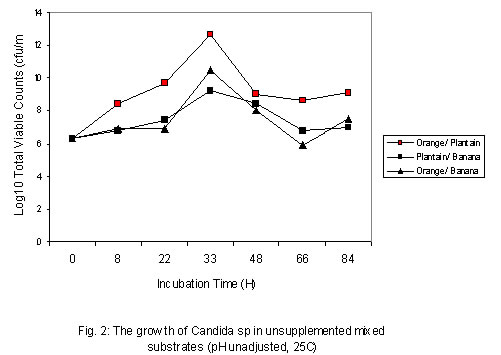
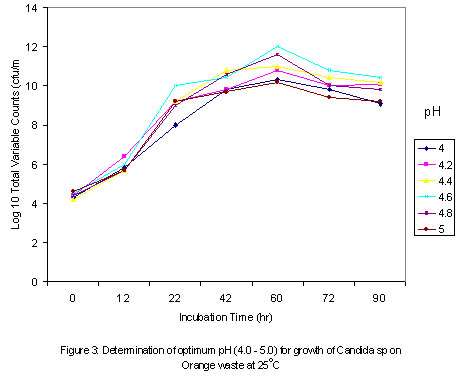
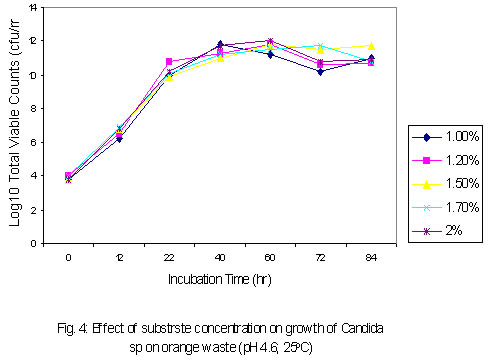
where The level of reducing sugars at different intervals of the incubation period was determined by aseptically withdrawing sample volumes (2ml) of the culture suspension into sets of clean centrifuge tubes and centrifuged at the high-speed setting using MSE model Minor 35 centrifuge to obtain clear supernatants. 1ml supernatant portions were then pipetted into clean test tubes and the amount of reducing sugars determined by the dinitrosalicyclic acid method described by Oso (1978). The level of total soluble carbohydrates were similarly determined on separate 1ml supernatant volumes as obtained above using the Anthrone procedure described by Allen et al. (1974). Yeast Cell Biomass Production: For this determination only supplemented dry ground orange wastes were used with unsupplemented media as controls. The media suspensions described above were supplemented with 0.025 - 0.20 % w/v dextrose and 0.1 - 0.8% w/v ammonium nitrate (NH4NO3) in duplicates. These were sterilized and inoculated as described previously. Inoculated flasks were incubated at room temperature on an orbital shaker for 60h. The solids fractions of the duplicate uninoculated flasks were harvested by centrifugation and supernatants decanted off. The harvested solids were then washed three times with sterile distilled water with centrifugation carried out intermittently between washings. The washed solids were then dried in a hot air oven at 80oC until constant weights were obtained. The difference between the weights of the inoculated and uninoculated flasks represented biomass yields. b) Comparison of biomass yields in different substrates Biomass production in supplemented and unsupplemented orange, plantain and banana waste media was compared. Media were prepared as described above and further steps in this determination were as previously described. RESULTS AND DISCUSSION Growth of the organism on potato dextrose agar (PDA) showed that it developed creamy-white, entire and raised colonies that were 1-2mm in diameter after 24h. Microscopic examination showed that the cells were Gram-positive, elongated and multiplied by bipolar budding (Table 1). Fermentation tests with the test strain yielded positive results with dextrose and fructose but negative for maltose, sucrose, mannitol and lactose (Table 1). Negative test was indicated by the absence of acid or gas production in the fermentation broth. Enzymatic studies for the production of amylases and cellulases showed that the test strain was unable to use starch and cellulose as sources of carbon and energy. Physiological tests showed that the test strain produced discrete colonies on PDA on incubation at room temperature. It however, failed to grow at 37ºC. One of the requirements of human microbial pathogens is that they must be capable of growth at 37ºC. This result presumably indicates the safety of the use of this organism for animal feed supplement production. An investigation of the amino acid requirements of this strain revealed that it was capable of growth on selective omission media lacking the amino acids screened (Table 1). This observation is in line with earlier studies by Snyder (1970), Dimmling and Seipenbusch (1978), Labaneiah et al. (1979) and Okada et al. (1980). The indication therefore is that the test strain was capable of synthesizing the amino acids and bases investigated, and could therefore be cultivated and harvested as a nutritional source of these amino acids in feed supplement production. Fermentation/Substrate Utilization Studies Comparison of growth in fruit wastes: A comparative study of growth levels of the production strain (Fig 1) in unbuffered suspensions of the fruit wastes revealed that highest growth, as log total viable count, was obtained with the orange waste (11.69) with slightly lower values for plantain (10.34) at 33h after incubation. The values for banana peels on the other hand were much lower (8.5). Further comparison of the growth levels in the three substrates was made using mixed substrate sources (Fig. 2). Results show that the growth level was highest (12.5) for the orange-plantain combination and slightly lower (11.7) for the plantain-banana combination. That for the orange banana mixture was approximately two log-cycles lower than the previous ones. The results also show that optimal growth was achieved after 33h. Further increase in numbers of viable cells was observed after 60h. This suggested the utilization of more than one substrate source of carbon and energy in the medium. The 33-h growth period contrasts with the 60h recorded when the medium was buffered (see Fig. 3). In terms of economics, with respect to time, the unbuffered could be assumed to be better as this fermentation option would provide for savings on energy costs, manpower hours and rate of production, i.e. turnover (Trilli, 1977; Moo-Young et al., 1981; and Gonzalez-Valdes and Moo-Young, 1981). Determination of optimum pH for growth - During growth in culture media one of the factors controlling the level of growth of any selected strain is the pH (Cruickshank et al., 1975). The pH would also determine the occurrence and types of particular contaminants in the culture medium. It is therefore necessary that the pH of growth of any selected strain be such that is optimal for this strain and at the same time reduces growth of contaminants. Preliminary investigations revealed that the test strain was capable of growth over a wide pH range of 3.0 to 6.2 (data not presented). The results also showed that optimum growth(s) could be achieved at pH 3.0, 3.4, 4.6 and 5.8. For further studies, the pH value of 4.6 was selected, the basis of which is presented in Fig. 3. Below a pH level of 4.6 (3.0 and 3.4) and above (5.8) mixing by rotary shaking was difficult as the media tended to be viscous. Since most saprophytic bacteria have wider pH growth range and can thrive at pH as low as 4.4 (Cruickshank et al., 1975), in the presence of these organisms, the optimum pH of 4.6 selected for the test strain has limited or reduced selective advantage. Lower growth pH within the range studied could be considered for large-scale cultivation of the test strain, as contamination may be more difficult to control at this level. This optimum poses the problem of controlling the observed increase in viscosity of media at the low pH values. This could be overcome by reducing the incubation period to 33 hours. Influence of Optimum Substrate Concentration - Results of tests with the production strain for the determination of optimum substrate concentration for growth as illustrated in Fig 4 show that growth, as total viable counts, was highest (12.1) for the substrate concentration of 2.0 % w/v. On the other hand the growth levels were approximately equal at substrate concentrations of 1.2 and 1.5 % (11.8) and least at 1.0% The results also show that for the substrate concentrations 1.2, 1.5, 1.7 and 2.0 % w/v the peak growth was obtained after 60h . Comparatively, for the 1.0- % concentration peak growth was observed after 40h. A probable reason for this observation is that at 1.0% concentration the problem of viscosity as reported earlier on is not existent and growth proceeds at a faster rate.
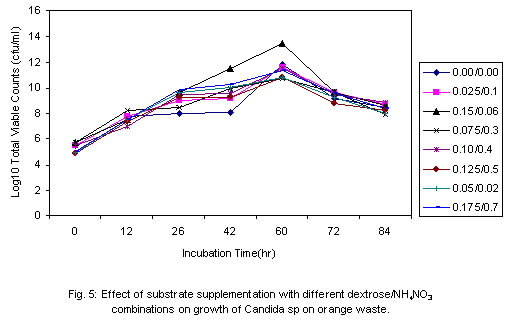
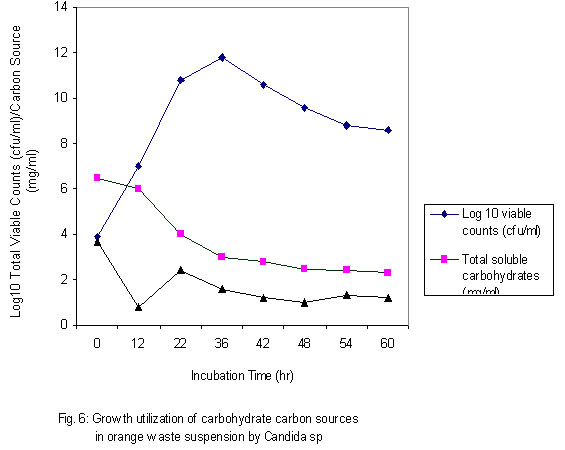
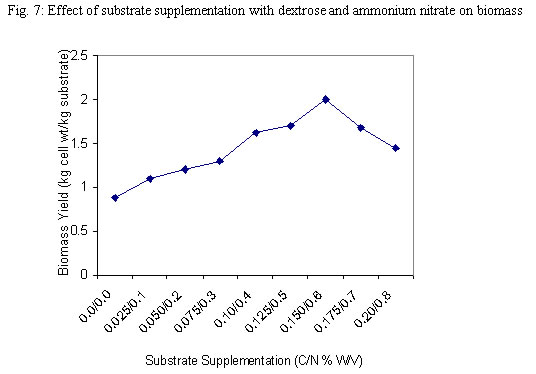
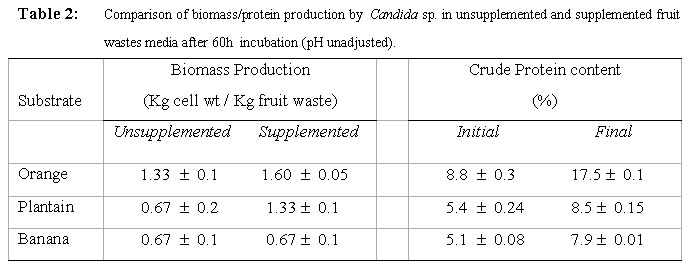
Although the best growth was obtained at substrate concentration of 2.0% w/v after 60h when only one growth cycle is considered, the effect of this relatively higher growth is lost after two log cycles when 1.5% concentration is considered at 84h. What is implied here is that the optimum concentration may not necessarily be translated as optimum in economic terms. As was observed during experiments to determine optimum pH for cultivation of the test strain, an increase in viscosity of the culture medium was noticed as the substrate concentration was progressively increased. The substrate concentration, and therefore medium viscosity, would influence the growth of the test strain in terms of agitation/aeration efficiency. Solomons (1983) has also reported that substrate concentration affects the yield of Saccharomyces cerevisiae when grown on an assimilable carbohydrate such as glucose or sucrose. For this organism, he reported that substrate levels in batch culture have to be kept very low, otherwise the substrate is oxidized to carbon dioxide and heat. He has similarly stated that this is one of the reasons why Candida species are preferred for single cell protein (SCP) production since they have a better regulation between anabolic and catabolic pathways, which prevent waste of substrates. This advantage could have contributed in part to the high yield recorded for the test strain. Influence of supplementation with carbon and nitrogen on growth of the test strain Preliminary investigations with the test strain (Adoki, 1987) revealed that this organism grew best in culture media supplemented with ammonium nitrate in contrast to supplementation with ammonium sulphate. Addition of phosphorus (KH2PO4) to media did not significantly improve growth. Further tests on the effects of mixed substrate supplementation on growth of the test strain were also carried out and results are presented in Fig. 5. The highest growth was recorded at the 0.15/0.6% w/v dextrose/ammonium nitrate combination, and this was more than one log cycle greater than the unsupplemented medium at peak growth point. On the other hand, the lowest growth was obtained with the 0.05/02% w/v combination. In all cases optimal growth was recorded after 60h incubation at room temperature and pH of 4.6. One criterion that is crucial in the selection of a yeast strain for protein production is the ability to grow in unsupplemented substrates or where supplementation is made it is simple and cheap. This criterion is satisfied with the results obtained where the test strain was found to grow well even in unsupplemented substrates with the exception of banana wastes. The higher level of added nitrogen indicates that the level of nitrogen in the growth substrates was more limiting than that of the sugars. Whilst supplementation with nitrogen source resulted in an increase in growth of the test strain, no such marked effect was observed when the production medium was supplemented with a source of phosphorus. This result could be explained in part by the relatively high levels of phosphorus in the substrates, and compared to carbon and nitrogen the level of phosphorus required by growing organisms is lower. The 0.2 to 0.4% recorded for the raw substrates (Adoki, 1987) also compares with that obtained for tropical citrus processing by-products by Crandal and Kseterson (1980). Results of investigations on the substrate conversion efficiency, as defined earlier on, of the test strain used for feed supplement production are presented in Fig. 6. The figure shows that at zero incubation time the concentration of total soluble carbohydrates and reducing sugars in the fruit wastes suspension growth medium were 6.5 and 3.7 mg/ml respectively. While there was no decrease in the level of soluble carbohydrates after an incubation time of 12h, a sharp decline in the level of reducing sugars was noticed for the same period. This indicated that the reducing sugars were more accessible to the test strain as immediate sources of carbon and energy compared to the other soluble carbohydrates. Based on the highest and lowest concentrations of soluble carbohydrates recorded during the incubation period, a utilization efficiency of 67.7% was obtained. Similarly, the efficiency obtained for reducing sugars was approximately 84%. This value also indicated greater utilization of the reducing sugars by the test strain. Figure 6 also illustrates the relationship between substrate utilization and cell growth. During the incubation period cell numbers reflected the sugar utilization patterns. Generally, a decrease in numbers of viable cells was recorded after 36h. This indicated that during this period, cell death was greater than cell growth. The substrate utilization efficiencies of 67.7% and 84% reported above were found to be higher than the values reported by Kamel (1979) for 4 yeast strains and are an indication of its potential for microbial biomass production. Yeast cell biomass is determined by the level of growth of the test strain, which in turn is ascertained by the log total viable counts. The effects of the substrate supplementation on biomass yields are presented in Fig. 7. The data suggest that yeast cell biomass production by the test strain was highest at the 0.15/0.6% (w/v) dextrose/ammonium nitrate supplementation level, giving a yield of about 2.03 kg/kg of supplemented orange waste. Comparatively, the yield for the unsupplemented substrate was 0.87 kg/kg of orange waste. At higher supplementation levels microbial biomass productions were found to be lower than that recorded at the 0.15/0.6% w/v supplementation level. The high yield recorded was similar to that reported by Okada et al. (1980) for Saccharomyces cerevisiae based also on the 0.15/0.6% supplementation level. On a comparative note also, results with supplemented substrates were similar to those reported by Moo-Young (1977) whose data suggested a yield of 0.85 kg/kg substrate. A comparison of biomass production in orange, banana and plantain wastes media as presented in Table 2 shows that production by the test strain was best in orange media under unsupplemented and supplemented conditions. Crude protein contents were also increased by about 100% in orange. Lower levels were recorded for plantain and banana. The level recorded for orange is comparable to that reported by Lequerica and Lafuente (1977). Conclusion: Investigations carried out during the study were primarily to determine the suitability of the chosen waste raw materials as substrates for the production of protein-rich feed supplements. The protein supplement derivable from the test strain could also be considered safe for use as animal and human feed supplement due to its inability to grow at temperatures beyond 25ºC. The low incubation temperatures also imply a savings on energy consumption and hence cost of production.The findings that the test strain was capable of growth on reducing sugars and some soluble carbohydrates imply that it could be used to reduce the levels of these carbon and energy sources in processing waste streams rich in them. Linked with the high biomass and protein yields recorded during this study, are the high substrate utilization or conversion efficiency and growth rates of the test strain. It could therefore be concluded that the process studied could be exploited commercially to produce protein-rich feed supplements. REFERENCES
Copyright 2002 - Journal of Applied Sciences & Environmental Management The following images related to this document are available:Photo images[ja02028f1.jpg] [ja02028t1.jpg] [ja02028f3.jpg] [ja02028f4.jpg] [ja02028f7.jpg] [ja02028t2.jpg] [ja02028f5.jpg] [ja02028f6.jpg] [ja02028f2.jpg] |
| |||||||||

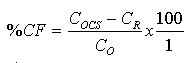
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}