 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
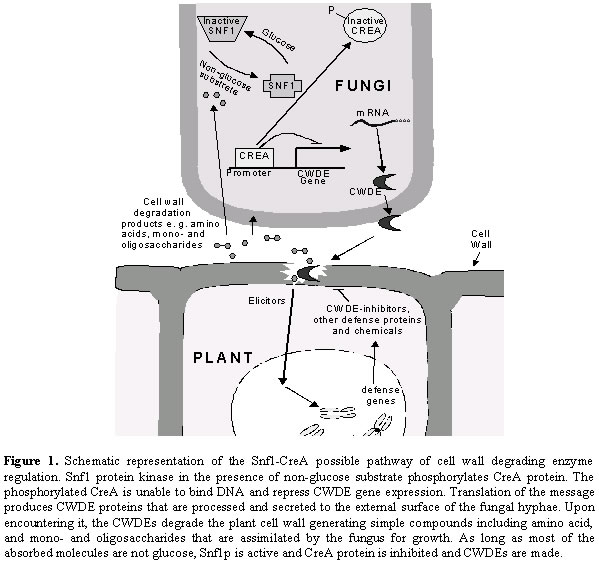
Journal of Applied Sciences & Environmental Management, Vol. 7, No. 1, June, 2003, pp. 5-8 Enzymes and fungal virulence TONUKARI, N J International Livestock Research Institute, P.O. Box 30709, Nairobi, Kenya. Tel: 254-2-630743, Fax: 254-2-631499, E-mail: J.TONUKARI@CGIAR.ORG Code Number: ja03001 ABSTRACT: This paper presents a comprehensive literature review of cell wall degrading enzymes (CWDEs). Plant pathogenic fungi secrete extracellular enzymes that are capable of degrading the cell walls of their host plants. These CWDEs may be necessary for penetration of the cell wall barrier, as well as for generation of simple molecules that can be assimilated for growth. Most of these enzymes are substrate-inducible and both specific and global regulators control their expression. CWDE-inhibitors have been isolated from plants and some evidence indicates that they are components of general resistance. @JASEM When microorganisms acquire the ability to enter and grow within healthy tissues, and subsequently circumvent plant host defense mechanisms, disease develops. Recent investigations on fungal pathogenicity have focused on defining the virulence factors produced by these pathogens, as well as the methods used to penetrate their hosts. Nevertheless, the ability of fungi to cause disease is dependent on their fundamental ability to penetrate and grow within plant tissues. Penetration into plant tissues and utilization of the nutrients found therein are requirements for all successful infections. Because cell wall constituents such as cellulose, xylan, pectin and proteins are typical among plants, the mechanism for penetration may be common across a variety of fungal pathogens. Most fungal pathogens and many non-pathogens produce large numbers of cell wall degrading enzymes (CWDEs) including pectinases, xylanases, cellulases, and proteases, which are capable of depolymerizing the various components of the host cell walls. Consequently, they have been studied in relation to tissue maceration, penetration, or nutrient acquisition from the cell wall polymers. In pathogenic fungi, these CWDEs may be required not only for tissue maceration, penetration, or nutrient acquisition, but for virulence as well. Fuel, Enzymes and Virulence A major reason for microbial attack on plants is to obtain nutrients for growth. To achieve this, they must first overcome the physical barrier presented by the plant cell wall. Fungi utilize the food substances in their immediate vicinity to promote growth toward other areas of food availability. This continuing need to reach new food supplies for growth and reproduction may be the most significant driving force of fungal virulence. Both the plant cell wall and the protoplasm contain nutrients that can be utilized by the growing fungus. Loosening and growing through the plant cell wall would allow the fungus access to simple sugars, amino acids, minerals and nucleotides that are abundant within the cytosol of the plant. In addition, degradation of the cell wall macromolecules would generate simpler compounds such as xylose, glucose, and amino acids that could be absorbed for growth. Fungal extracellular enzymes may have the potential to degrade the structural cell wall constituents of living plants, but proving that such interactions initiate or promote pathogenesis has been difficult due to redundancy of these enzymes. For example, C. carbonum secretes at least five enzymes that can degrade the xylan component of the maize cell walls (Tonukari et al., 2002). Functional redundancy provides a means to adapt to different conditions, and is indicative of processes with vital importance to an organism. Despite this redundancy, however, single constitutive pectinase genes have been shown to contribute to the virulence of Aspergillus flavus on cotton bolls (Shieh et al., 1997) and Botrytis cinerea on tomato (ten Have et al., 1998). While a specific CWDE may be very important for virulence in one pathogenic fungus, others may likely require a combination of several CWDEs (Tonukari et al., 2000). CWDE regulation The expression of most CWDEs depends on external conditions such as substrate availability or the specific stage of disease development. The majority of CWDEs are made at low basal levels on sugars such as glucose or sucrose, but are highly expressed when the fungus is grown on the appropriate substrate. For example, the expression of C. carbonum xylan-degrading enzymes are induced by xylan, and inhibited by glucose (Tonukari et al., 2002). Kolattukudy et al. (1995) also reported that a cutinase present in spores of F. solani pisi releases small amounts of cutin monomers upon contact with the plant surface, which then trigger cutinase gene expression. This induction of CWDEs by cell wall components suggests a possible signaling pathway from the plant cell leading to the expression of CWDE-encoding genes inside the fungal cell. Investigations have indicated that both specific and global regulators control the expression of CWDEs. XlnRp, a transcriptional activator of the xylanolytic system, has been identified in A. niger (van Peij et al., 1998). While Chaetomium gracile xylanase A gene (cgxA) is repressed by binding of a protein designated AnRP (Mimura et al., 1999). The involvement of CreA protein, the carbon catabolite repressor, in the regulation of CWDEs has been reported in several fungi. CreA binds to the promoter region of several genes and inhibits their expression (Ronne, 1995). The gene encoding the CreA has been identified in Trichoderma reesei, C. carbonum, and other fungi (Mach et al., 1996; Tonukari et al., 2003). Inhibition of gene expression by the CreA complex has been postulated to be relieved a protein kinase, Snf1p whose gene has been isolated in C. carbonum (Tonukari et al., 2000). Therefore, a gene disruption that abolishes the Snf1 activity should down-regulate CWDE expression. Mutation of the SNF1 gene in C. carbonum leads to a decreased expression and activities of several CWDEs including β-1,3-glucanases, pectinases and xylanases (Tonukari et al., 2000). The C. carbonum snf1 mutant also shows reduced virulence on maize, which is consistent with the hypothesis that CWDEs are virulence factors. plant response to CWDEs Although plants are exposed to many microorganisms, some of which are potential pathogens, they are naturally resistant to the vast majority. In addition to the cell wall barrier, other pathways that activate a battery of defense mechanisms against potential pathogens may also exist. In general, resistance could involve small molecules or proteins that inhibit or promote the digestion of the fungal secreted enzymes thereby denying access to the host cell. For a pathogen to be successful, it must be able to circumvent or overcome these antifungal defenses. The identification of CWDE inhibitors in plants provides indirect evidence that CWDEs are virulence factors. A glycosylated, basic protein that inhibits A. niger and Trichoderma viride endo-1,4-β-xylanases has also been identified in wheat (McLauchlan et al., 1999). Furthermore, an inhibitor of pectin methylesterase from kiwi fruit is also a glycoprotein, but has an acidic isoelectric point (Giovane et al., 1995). Polygalacturonase-inhibiting proteins (PGIP) are typically effective against fungal endopolygalacturonases (Cervone et al., 1989). The PGIPs are predominantly bound to plant cell walls, and their levels increase in bean hypocotyls when primary leaves begin to develop (Salvi et al., 1990). PGIP forms specific and reversible high affinity complexes with fungal polygalacturonases, thereby regulating the activity of the fungal enzymes (Cervone et al., 1989). High-level expression of pear PGIPs in transgenic tomato fruits leads to increased resistance to B. cinerea (Powell et al., 1994). The PGIP inhibition of B. cinerea polygalacturonases slows the expansion of disease lesions and the associated tissue maceration in transgenic tomato plants (Powell et al., 2000). A schematic representation of the expression of cell wall degrading enzyme in fungi and its interaction with plant defense mechanism is depicted in Figure 1. It is envisaged that more CWDE inhibitor proteins will be identified in the near future. Conclution Virtually all fungi produce a great abundance and variety of CWDEs that may be needed for softening up of the plant cell walls for penetration by fungal hyphae, as well as provision of nutrients for growth. The application of molecular technologies to the study of fungal-plant interactions offers a new and more definitive approach for examining the role played by CWDEs in disease development. Resistance to CWDEs may be part of the constitutive and/or acquired resistance that plants possesses against various pathogens. Any factor that will inhibit the production of CWDEs in fungi could lead to reduction in virulence. REFRENCES
Copyright 2003 - Journal of Applied Sciences & Environmental Management The following images related to this document are available:Photo images[ja03001f1.jpg] [ja03001t1.jpg] |
| |||||||||

{kind=link}
{kind=link}