
|
Journal of Applied Sciences and Environmental Management
World Bank assisted National Agricultural Research Project (NARP) - University of Port Harcourt
ISSN: 1119-8362
Vol. 8, Num. 2, 2004, pp. 39-44
|
Journal of Applied Sciences & Environmental Management, Vol. 8, No. 2,
Dec, 2004, pp. 39-44
Determination of the infection densities of mudfish Eustrongylides in Clarias
gariepinus and C. anguillaris from Bida floodplain of
Nigeria
1* IBIWOYE, T I I.; 2BALOGUN,
A M; 3OGUNSUSI, R A; 1AGBONTALE, J J
1Pathobiology Programme, Artisanal Fisheries Division,National
Institute for Freshwater Fisheries Research (NIFFR),P. M. B. 6006, New Bussa,
Niger State, Nigeria.
2Department
of
Fisheries and Wildlife,Federal University of
Technology,
P.M.B. 704, Akure,Ondo State, Nigeria.
3Department
of
Animal Production and Health,Federal University of
Technology,
P.M.B. 704, Akure,Ondo State, Nigeria.
*Corresponding author
Code Number: ja04022
ABSTRACT
The
infection densities of Eustrongylides africanus larvae were analysed
according to the season, sex and distribution. Its prevalence, intensity and
abundance
was 26.5 ± 15.4%, 2.1 ± 0.9 and 0.7 ± 0.5 worms per fish, respectively for the
wet season and the dry season had 32.9 ± 5.4%, 2.5 ± 0.4 and 0.9 ± 0.2 worms
per fish, respectively. These incidences increased greatly during the dry season,
being higher in the females than the males for both seasons of the year. Its
lifecycle is complex and indirect with fish as intermediate, reservoir (alternative)
or final (definitive) hosts. However, this is the first report and record of
the incidence and lifecycle of infections with larvae of Eustrongylides africanus in
free catches freshwater mudfish Clarias from Bida floodplain of Nigeria. @JASEM
The
occurrences of helminth parasites in fishes have been extensively studied in
various water bodies in Nigeria.
Some works included those done on the River Niger at Shagunu; River Niger at
Kainji Reservoir, Benue and Ogun Rivers; on the Coast of Lagos; on the Cross
River estuary; on the Niger Delta Area; on the Lake Chad; from Zaria (Aken’ova,
1999; Auta et al., 1999); on the Imo River; from the Anambra River Basin
(Ezenwaji and
Ilozumba, 1992); Ile-Ife;
from the Jos Plateau (Anosike et al., 1992; Omoregie et al., 1995);
on the Opi Lake; on the River Kaduna (Emere, 2000) and on the Kandole Shela Stream
in Sokoto. Based on these criteria (morphological, anatomical
and colour description) as well as on ecological data over ten species
of Clarias have been identified in West Africa (Sydenham,
1980). No studies, however, have been made concerning the quantitative assessment
of worm
burdens in free catches Clarias from Bida floodplain of Nigeria.
This
study was undertaken to determine the infection densities of Eustrongylides africanus larvae
in Clarias from Bida floodplain, according to season and sex, for the
first
time.
MATERIALS
AND METHODS
The study covered an
area located between longitude 5o 45’ to 6o 15’E and
latitude 8o 30’ to 9o 10’N within the southern Guinea
Savannah zone of Nigeria (Areola et al.,
1992). 2700 Clarias of different sizes, sexes, lengths and weights randomly
selected were 5-10% of the catches from four sampled locations (Doko, Dokoji,
Fokpo and Dutsu) in the Bida floodplain. They were handled in humane manner,
thoroughly examined and dissected to recover helminth larvae, for direct numerical
count using a tally counter, and the sites of infection recorded at NIFFR Fish
Health Diagnostic Laboratory. Its prevalence, intensity and abundance were determined
for sex and seasons of the year as described by Margolis et al. (1982).
RESULTS
AND DISCUSSIONS
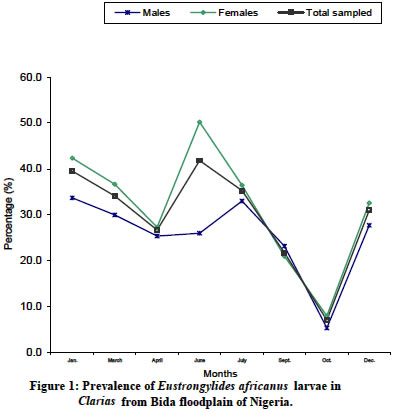
The prevalence of E. africanus larvae
infection in Clarias from Bida floodplain is shown in Figure
1. The
males’ prevalence of 33.7% in January decreased to 26% in June, and increased
to its peak of 33% in July followed by sharp fall to lowest value of 5.4% in
October
with a sharp rise to 27.7% in December. The females’ prevalence of 42.4% in January
decreased to 27.3% in April followed by a sharp rise to its peak of 50.2% in
June, also with a sharp fall to its lowest value of 8.0% in October later by
a sharp rise to 32.6% in December. The prevalence for the total sampled of 39.6%
in January decreased to 26.7% in April followed by a sharp rise to its peak of
41.9% in June followed by a sharp fall to its lowest value of 7.1% in October
later by a sharp rise to 31.1% in December. Both the females and total sampled
had their peaks in June while the males had their peak in July. The trend of
prevalence
is about the same for the males, females and total sampled since all of them
had falls in April and October. The
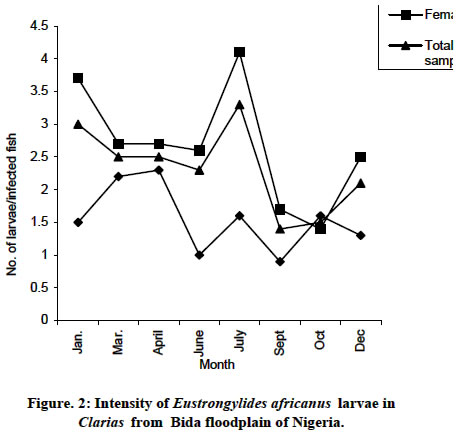
intensity of Eustrongylides africanus larvae in Clarias of
Bida floodplain is shown on Fig 2. The males intensity of 1.5 larvae per infected
fish increased to 2.3 in April followed as sharp fall to 1.0 in June with characteristic
rises and falls between July and
to 1.3 in December.
The females’ intensity of 3.7 larvae per fish gradually decreased between March
and June to 2.6 followed a sharp rise to its peak of 4.1 in July also followed
by a sharp fall to 1.4 in October and later by a sharp rise to 2.5 in December.
The intensity for the total sampled of 3.0 larvae per fish in January gradually
decreased between May and April to 2.7 followed by a gradual rise to its peak
of 3.5 in July with a sharp fall to its lowest value of 1.4 in September and
followed by a rise to 2.1 in December. All
of them had peaks and falls in July and September, respectively. The trends
of intensity
are about the same for the females and total sampled but contrarily for the
males. The abundance of Eustrongylides africanus larvae infection
in Clarias from
Bida floodplain is shown in Figure 3. The
males’ abundance of 0.5 per sampled fish in January increased to its highest
peak of 0.7 in March, decreased to 0.3 in June and rose to 0.5 in July followed
by a fall to its lowest value of 0.1 in October and later rose to 0.4 in December.
The females’ abundance of 1.6 larvae per sampled fish in January decreased sharply
to 0.7 in April, rose sharply to 1.5 in July, fell sharply to 0.1 in October
and later rose sharply to 0.8 in December. The abundance for the total sampled
of 1.2 larvae per sampled fish in January fell sharply to 0.7 in April, rose
sharply to 1.2 in July and fell sharply to 0.1 in October and later rose sharply
to 0.7 in December. All of them had peaks
in July and falls in October. The trends of abundance are about same for the
females
and total sampled but contrarily for the males.
It is evident
that female fishes are more frequently infected with parasites than the males
from the findings of this study. This observation agreed well with the findings
of Mhaisen et al. (1988) that female fishes were generally more liable
than males to infections with cestode, nematode, acanthocephalan, crustacean
and copepod parasites. The mean prevalence,
intensity and abundance of Eustrongylides africanus larvae in Clarias for
the dry season is higher than the wet season of the year. Fishes are susceptible
to heavy infestation with parasites mainly in the early rain when fishes are
weakened by hibernation (a state of exhaustion). Larger fishes are heavily
parasites than smaller ones. The intensity and prevalence of parasites infection
increases
with increasing length, size and age of the fish host. And that the food amount
contained in the intermediate host of the parasite might cause the increase
in infection level.
Eustrongylides species
have complex, indirect life cycle (Yanong, 2002) with fish as intermediate,
paratenic (alternative) or final (definitive)
hosts. Where the fish is the intermediate host, the nematode eggs/larvae
enter an invertebrate or a fish intermediate host prior to being eaten by
or entering
the final host. The final host (which contains the reproductive adult stage
of the nematode may be a piscivorous (fish – eating) fish, mammal, amphibian,
reptile or bird. Where the nematode has the ability to survive in “alternative” organisms,
known as “paratenic” or reservoir hosts, which are not required for completion
of the life cycle, but they can contain infective nematode life stages and
be a source of infection. They can be fish, worms or other aquatic organisms
that can eat the nematode eggs/larvae. Where the fish is the final host,
the nematode eggs/larvae enter an aquatic invertebrate intermediate host
such as
a copepod, annelid worm or insect larva prior to being eaten by or entering
the final host fish. The eggs of all Eustrongylides species
are very tough and can
easily survive for some time in fishponds. At about 77°F, it can
take anytime from 3 – 4 1/2 months from the time fish become infected. This
means that, after sterilization of ponds, if fish – eating birds do infect
the ponds with Eustrongylides eggs, the producer may not see a problem
until harvesting the fish, 3 – 4 months later, as this is approximately the
time required for the eggs to hatch and become the L3 stage which infects the
fish. After this
3 – 4 months period, fish raised on ponds
with a population of fish – eating birds have a greater chance of becoming
infected as the number of nematode increase overtime. Adults are found in fish – eating
birds, the eggs are shed by the aquatic birds into ponds, where they develop
into a life stage that is consumed by an oligochaetes or annelid worms where
it develops further into a third large stage “L3” which can infect fish when
eaten. Once the annelid worms containing the L3 stages is eaten by a fish and
digested, the nematodes migrate (within the fish) into the body cavity and,
frequency, over the external surface of internal organs such as the liver.
Some recent studies,
however, suggest that E. ignotus, commonly found in mosquito
fish, which is a
close relative of Eustrongylides africanus, may be able to complete
its life cycle
without the need for a tubifex worm (Coyner et al., 2002). The first
intermediate hosts are oligochaetes or annelid worms (Shah-Fischer and Say,
1989). Complementary
(reservoir, alternate, paratenic) hosts are fish, where encysted larvae are
found. The death of the host might stimulate the nematodes to emerge from their
cysts,
migrate through the body wall to the surface of their dead host and may survive
for over 24hrs when a dampened condition exists. The specimens of fourth stage
of Eustrongylides species were morphologically
consistent with materials described by Lichtenfels
and Pilitt (1986). Adult Eustrongylides africanus (Jaegerskiold,
1909), the only species so far reported from Africa was found and described
only from the proventriculus
and stomach walls of the female cormorants Phalacrocorax africanus (Gmelin)
and
in a wide diversity of other aquatic fish-eating birds (definite  hosts). Eustrongylides are
known from a diversity of fishes, mammals, amphibians, reptiles and birds (Hoffmann,
1999) and their relatively great
abundance suggests they are common in the habitats where fish forage. But exceptionally
rare parasites of carnivorous mammals associated with riparian or lacustrine
habitats. The prevalence recorded in this study may
indicate that larvae are particularly abundant in fishes and amphibians available
to Rivers Kaduna and Niger and other tributaries supplying water to Bida floodplain;
consistent with the potential that humans
could be exposed to infection. hosts). Eustrongylides are
known from a diversity of fishes, mammals, amphibians, reptiles and birds (Hoffmann,
1999) and their relatively great
abundance suggests they are common in the habitats where fish forage. But exceptionally
rare parasites of carnivorous mammals associated with riparian or lacustrine
habitats. The prevalence recorded in this study may
indicate that larvae are particularly abundant in fishes and amphibians available
to Rivers Kaduna and Niger and other tributaries supplying water to Bida floodplain;
consistent with the potential that humans
could be exposed to infection.
All
evidences indicated that the larva is Eustrongylides africanus and
that a wide diversity of aquatic fish-eating birds, which are extremely numerous
along the floodplain serve as final hosts with tilapias their predominant food.
Adult Bagrus and Clarias are too large to be eaten by aquatic
fish-eating birds or any other potential hosts of Eustrongylides. The
infection to the final host is apparently transmitted mainly or exclusively
through tilapias. Bagrus and Clarias apparently become infected
with larval Eustrongylides by ingesting infected tilapias a dominant
food item in their diets. The ability of Eustrongylides larvae to re-establish
in a new intermediate host was experimentally
demonstrated with E. ignotus. Adult E.
africanus are apparently not particularly host specific since they have been
reported from several species of aquatic birds widely distributed on the African
continent, e.g. cormorants Phalacrocorax africanus (Gmelin), herons Ardea goliath (Cretzschmer),
snake birds Anhinga rufa (Lacepede), pelicans Pelecanus rufescens (Gmelin)
and spoon bill Platalea leucordia (Linnaeus) (Jaegerskiold, 1909). Eustrongylides species can be found within the body
cavity, encapsulated on the liver and other organs, but outside the intestinal
tract of fish. Eustrongylid nematodes can affect a number of different species,
including yellow perch, pumpkin seed, mummichug, guppies, gar, danios and angelfish.
Affected fish typically have bloated abdomens (dropsy), as the nematodes frequently
migrate into body cavity and can be quite large. Eustrongylids are typically
very long, coiled and red (due to the presence of haemoglobin) and an infected
fish often has more than one nematode in its body cavity. If a
feeder fish containing Eustrongylides species are fed to other fish, the nematode can migrate
out of the feeder fish and into the muscles or other organs of the fish that
just consumed them. After migrating into the muscle, this nematode can cause
lesions that look superficially similar to a grub. Infection would tend to build
up in already infected environment due to re-infected as this worm is viviparous
and since contact with other
infected fish could be less in the river with the large volume of water than
in the floodplains or fish ponds with smaller water volume. The differences in
size of nematode larvae might have resulted
from the differences in the site of infection. Most piscivorous birds are unlikely
to find Clarias easy to handle because of its pectoral spines so encystment
of this
larva on Clarias might prove to be a dead – end in most cases.
Figure 4
ACKNOWLEDGEMENT
The authors are grateful to the
Federal Government of Nigeria through the Federal Ministry of Agriculture
and Rural development and National Institute for Freshwater Fisheries Research
(NIFFR) New Bussa for providing the fund for this
study.
REFERENCES
-
Aken’ ova, T. O (1999). Helminth
infection of the gills of Clarias species in Zaria Nig. J.
Parasitol. 20: 113-121..
-
Anosike, J. C., E. Omoregie; P. C.
Ofojekwu and I. E. Nweke
(1992). A survey of helminth
parasites of Clarias gariepinus in Plateau State, Nigeria. J.
Aquat. Sci. 7: 39-43.
-
Areola, O ; O. Irueghe; K. Ahmed;
B. Adeleke and G. C. Leong
(1992). Certificate physical and
human geography for secondary schools. University Press Plc, Ibadan. 406pp.
-
Auta, J; S. J. Oniye and J. A. Adakole
(1999). The helminth
parasites of the gastro-intestinal tract of Synodontis species in Zaria,
Nigeria. Zuma JPAS2 (2): 47-53.
-
Coyner,
D.F. ; M.G. Spalding and D.J. Forrester (2002). Epizootiology
of Eustrongylides ignotus in Florida: distribution, density and
natural infections in intermediate hosts. J. Wildlife Dis. 38 (3):
483 – 499.
-
Emere, M. C (2000). Parasitic infection
of the Nile perch Lates niloticus (L) in River Kaduna. J. Aquat. Sci. 15: 51-54.
-
Ezenwaji, H. M. G and P. C. O. Ilozumba
(1992). Helminth fauna of four West African small Clarias species
(Osteichthyes: Clariidae) from
Nigeria. J. Afr.Zool. 106: 391 – 400.
-
Hoffmann, G. L (1999). Parasites of North American Freshwater
Fishes second edition. Comstock
Publishing Associates, Ithaca, N.Y. 208pp.
- Ibiwoye, T. I. I; A. N. Okaeme; A.
M. Balogun and R. A. Ogunsusi
(2000). Updating the helmith parasites fauna of freshwater fishes in Nigeria
in the new millennium: First occurrence of Eustrongylides africanus (Khalil & Thurston,
1973) larvae in Clarias species of
Nigeria. 15th Annual Conference of the Fisheries Society of Nigeria
(FISON), Jos,
Plateau State. 19th – 24th march, 2000.
-
Jaegerskiold, L. A (1909). Nematoden
aus Aegypten und dem Sudan. Results of the Swedish Zoological Expediction
to Egypt and the White Nile 1901 (3):
1 – 66.
-
Khalil, L. F (1999). Confirmation
of the helminth parasites in Clarias species from Bida floodplain.
Personal
communication.
-
Lichtenfels, J. R and P. Pilitt
(1986). Eustrongylides sp. (Nematoda: Dioctophymatoidea): Differentiation
of third and fourth-stage larvae from Killifish, Fundulus sp.
Collected in Chesapaeke
Bay area, U.S.A. Proceedings of Helminthological Society of Washington.
53: 144 – 148.
-
Margolis, L.; G. W. Esch; J.C. Holmes;
A. M. Kuris and G. A. Schad (1982). The use of ecological terms in parasitology
(report of an
ad hoc committee of the American Society of Parasitologists). J. Parasitol. 68:131 – 133.
-
Mhaisen, F. T; N. K. Al-Salim
and N. R. Khamees (1988). Occurrence of parasites of the freshwater
mugilid fish Liza abu (Heckel) from Basrah, southern
Iraq. J. Fish Biol. 32: 525 – 532.
-
Omoregie, E; E. B. C. Ufodike and
C. O. E. Onwuliri (1995). A comparative survey of lminth parasites of
the Nile Tilapia, Oreochromis niloticus from a fish farm and petroleum
polluted
freshwater. J.
Aquat. Sci.10:
7-14.
-
Paperna, I (1996). Parasites, infections
and diseases of fishes in Africa: An update. CIFA Technical Paper. 31:
157 – 170.
-
Shah-Fischer, M and R. R. Say (1989). Manual
of tropical veterinary parasitology. CAB International, Wallingford,
Oxon, U.K. 473p
-
Sydenham, D. H. J (1980). New species
of Clarias from West Africa (Pisces: Clariidae) Rev. Zool. Afr. 93: 659 - 677.
-
Yanong, R.P.E. (2002). Nematode (Roundworm) infections in fish. First
edition. Cooperative Extension Service, Institute of Food and Agricultural
Sciences,
University of Florida, U.S.A. 16pp.
Copyright 2004 - Journal of Applied Sciences & Environmental Management
The following images related to this document are available:
Photo images
[ja04022f2.jpg]
[ja04022f4.jpg]
[ja04022f1.jpg]
[ja04022f3.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}