 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Applied Sciences & Environmental Management, Vol. 9, No. 1, 2005, pp. 25-30 Activity of Zymomonas species in palm-sap obtained from three areas in Edo State, Nigeria. OBIRE, O. Department of Applied and Environmental Biology, Rivers State University of Science and Technology, Nkpolu-Oroworukwo, P.M.B. 5080, Port Harcourt. Code Number: ja05005 ABSTRACT:The bacterium Zymomonas mobilis was isolated from fresh palm-sap samples from three different locations as to determine the contribution of the bacterial isolate to alcohol production in palm-wine. Carbohydrate (sugar) fermentation, alcohol tolerance, and growth of the bacterium at different pH and temperature values were determined; and a comparison of sugar fermentation by Z. mobilis and Saccharomyces cerevisiae was also determined. Z. mobilis contributes much to the fermentation of palm-wine due to its ability to ferment sucrose, glucose and fructose which are the main sugars in palm-sap. The bacterium reached its maximum density of 2.72 x 107 cells/ml between 12 and 15 hrs after tapping of the palm-sap. Z. mobilis is acid and alcohol tolerant being able to grow at pH values between 4.0 and 7.0 and in alcohol concentration of between 2.5% and 15% respectively. Z. mobilis was also found to ferment sugars at a faster rate than Saccharomyces cerevisiae. Statistical analysis of the data obtained using ANOVA showed that there is no significant difference at p = 0.05 level of significance in the growth of isolates of Z. mobilis obtained from the three locations under different pH, alcohol and temperature values. @JASEM
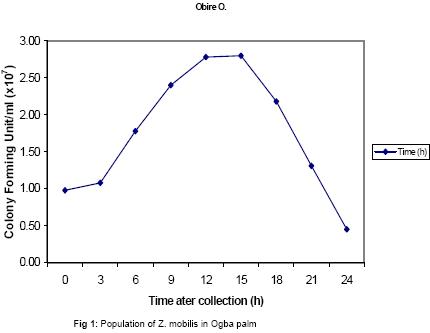
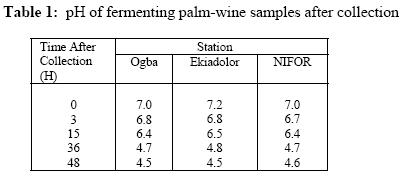
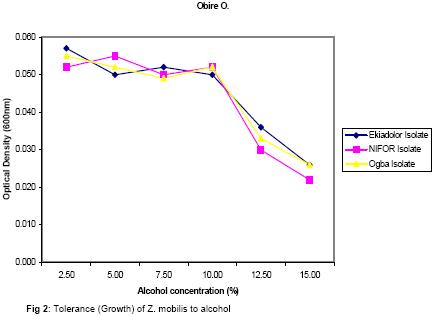
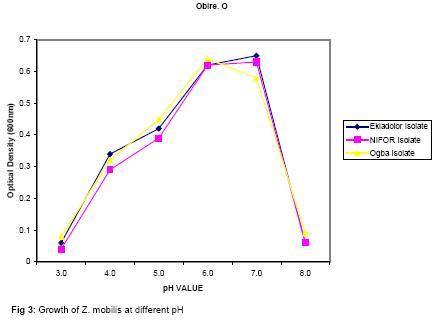
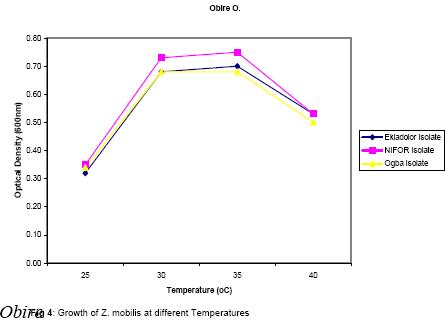
The presence of various microorganisms especially the bacteria and yeasts responsible for the fermentation of palm-wine has been reported by Bassir (1962), Faparusi (1966) and Okafor (1977). During fermentation, the sugars in the palm-sap are metabolized to alcohol and organic acids with the result that the sap loses its sweetness (Okafor, 1975). The types of bacteria present appear to depend on the stage of fermentation and the composition of the sap (Bassir, 1962; Okafor, 1977). Although alcohol production is common among yeasts, it is rare among bacteria (Ingraham and Ingraham, 2004). Yeasts are used to make most alcohol beverages. But pulque is an exception. Pulque is an alcoholic beverage from the juice of the agave plant fermented by Zymomonas mobilis (Talaro and Talaro, 1999; Ingraham and Ingraham, 2004). In Mexico, the distilled spirit tequila is traditionally made from the fermentation of juices from the agave plant using the bacterium Z. mobilis (Ingraham and Ingraham, 2004; Nester et al., 2004). Zymomonas species are perhaps the most important alcoholic fermenters of the bacterial group in plant saps and juices. Zymomonas are facultative anaerobes with both respiratory and fermenting capabilities. As in yeast, Zymomonas decarboxylates pyruvate non oxidatively with the formation of acetaldehyde; which is subsequently reduced to ethanol by the Entner-Doudoroff’s pathway (Swings and De Ley, 1977; Talaro and Talaro, 1999; Ingraham and Ingraham, 2004). The production of acetaldehyde and the characteristic fruity odour of Zymomonas also contribute to the odour and taste of wines (Swings and De Ley, 1977; Ingraham and Ingraham, 2004). The difficulty of storing palm-wine to retain its normal characteristics due to the fermentative ability of probably Zymomonas species and other microorganisms present in the wine has been a major problem in the bottling of palm-wine in Nigeria and consequently its distribution for consumption. In addition to their fermentative ability, Zymomonas species are known to be harmless to man and cattle. Some of its fermentation products are found useful in the treatment of various diseases ranging from chronic enteric metabolic disorders to gynecological infections (Swings and De Ley, 1977). The presence of Zymomonas species in palm-wine may thus be beneficial to man. The present study therefore is aimed at the isolation and identification of Zymomonas species in palm-sap and to ascertain whether the species is present in palm-sap from different locations. Attempts shall also be made to determine at what stage of fermentation the bacterium is most abundant in the palm sap and to determine the contribution of the bacterium to the fermentation of sugars in palm-sap and therefore to alcohol production in palm-wine. MATERIALS AND METHOD Palm-wine sample were collected from Elaeis guineensis by the method of “inflorescence tapping” as described by Faparusi (1966) and Okafor (1972). The palm-wine samples were simultaneously tapped from three different ecological areas (Ogba, Ekiadolor and the Nigeria Institute for Oil Palm Research, NIFOR) in Edo State of Nigeria. The palm-wine was put in sterile plastic containers and the containers were immediately immersed in freezing mixture of sodium chloride and ice chips and then taken to the laboratory for analysis. This method of collection is known to reduce fermentation rate considerably (Bassir, 1962). Media with the following compositions, as prescribed by swings and De Ley (1977), were used for the detection, isolation and enumeration of Zymomonas species from palm-wine: Detection medium: Malt extract 3.0g, Yeast extract 3.0g, Glucose 20g, Peptone 5.0g, Ethanol 30ml, Distilled water 1 litre, pH 4.0; Isolation and Enumeration Media: A. (i) Standard Solid Media Yeast Extract 5.0g, Glucose 20g, Agar 20g, Distilled water 1 litre, pH 6.8, (ii) MYPG Medium Malt extract 3.0g, Yeast extract 3.0g, Peptone 5.0g, Glucose 20g, Agar 20g, Distilled water 1 litre, pH 4.8, (iii) P.G.A. Medium Peptone 5.0g, Glucose 20g, Agar 20g, Distilled water 1 litre, pH 6.4, B. Standard liquid medium Yeast extract 5.0g, Glucose 20g, Distilled water 1 litre, pH 6.8, The pH of each medium was adjusted to the appropriate pH value using either NaOH or HCI. Each medium was treated with actidione (Cycloheximide) to inhibit yeast growth, and then autoclaved at 121°C for 15 minutes. Population of Zymomonas in palm-sap samples: The solid media were separately used for the spread plate technique in counting Zymomonas colonies. Plates inoculated with palm-sap samples were incubated in an anaerobic jar in which Gas Pak Sachet was placed to exhaust the oxygen in the jar and produce carbon dioxide saturated atmosphere. Incubation was usually for 2 days at 30°C after which the number of Zymomonas colonies on the plates was counted. Enumeration was carried out at 3 hr intervals to establish when the Zymomonas population reached their maximum density. Identification of Zymomonas sp: Colonies suspected to be those of Zymomonas were isolated from the spread plates and purified by streaking on freshly prepared media and incubated for 2 days at 30°C in the anaerobic jar. Isolates from such fresh plates were subjected to the following tests for the characterization of the isolates: Gram stain, catalase, motility, oxidase, urease, nitrate reduction, indole, carbohydrate fermentation and tolerance to alcohol tests. Carbohydrate (sugar) fermentation: The ability of the bacterium to ferment various carbohydrates using glucose, fructose, sucrose, maltose, lactose and arabinose was determined by growing the isolate in liquid standard medium containing 1% (w/v) of the particular carbohydrate. Durham-tubes were inverted into the culture tubes for gas collection. Incubation was at 30°C for 24hr. and uninoculated broths were used as control. Determination of alcohol tolerance: The alcohol tolerance of the isolate was determined by culturing it in media containing 2.5%, 5%, 7.5%, 10%, 12% and 15% ethanol. The alcohol was aseptically added to sterile medium. The cultures were incubated at 30°C for 24 hrs. The absorbance of the culture were recorded at 600nm using a Busch and Lomb Spectronic 20. Uninoculated broth was used as the control. Growth of isolate at different pH and Temperature: The growth of the isolate was examined at pH 3.0 to 8.0 using the standard broth. The growth was measured using a Spectronic 20 at 600nm. The Spectronic 20 was standardized with uninoculated broth. In addition, changes in pH value during fermentation were monitored at intervals of 0h, 3h, 15h, 36h, and 48h after collection. The following temperatures 25o, 30o, 35o, and 40°C were used to study the growth rate of the isolate for 24hrs. The absorbance at 600nm was recorded. Fermentation of sugar by Zymomonas mobilis and Saccharomyces cerevisiae: Two present (2%) solution of sucrose was used in this experiment. Nine milliliters of the sucrose solution was dispended into screw-capped test tubes with Durham tubes inverted into them to collect the gas produced. The test tubes were divided into six groups of four tubes each. Individual test tube in each group was inoculated as follows: Group A: 1 ml suspension of Z. mobilis , Group B: 1 ml palm-wine, Group C: 0.5 ml Z. mobilis and 0.5 ml palm-wine, Group D: 1 ml S. cerevisiae, Group E: 0.5ml Z. mobilis and 0.5ml S. cerevisiae, Group F: 0.5ml S. cerevisiae and 0.5 ml palm-wine. The tubes were incubated at 30°C for 24hrs after which, the volume of gas produced by the different groups were compared. RESULTS AND DISCUSSION Zymomonas mobilis was identified as the only Zymomonas species present in palm-wine obtained from the three different ecological sites. It was Gram negative, catalase positive, anaerobic, plump rods with an unusual width. It was heterofermentative, producing gas from glucose, fructose and sucrose. Maltose and arabinose were not fermented. All the isolates in this study were motile, but oxidase and urease negative. These observations were similar to those reported by Swings and De Ley (1977) on Z. mobilis isolated from various sources. Z. anaerobia also has similar characteristics with Z. mobilis except that the former is non-motile. The population of the bacterium was almost the same for all samples and ranged from 98 x 105 cells/ml in Ekiadolor and Ogba samples to 99 x 105 cells/ml for the NIFOR sample on arrival in the Laboratory. No work has been reported on the count of Z. mobilis in palm-wine. Okafor (1975) did some work on the organism but did not determine its population. The population of Z. mobilisin the Ogba palm-wine used in this study is shown in Fig. 1. The organism reached its maximum density between 12 to 15hrs after collection. The initial density was 98 x 105 cells/ml. This reached the maximum of 2.72 x 107 cells/ml after 12h and decreased after 15hr until the density after 24hrs was almost 50% of the initial density. The initial rise in density within the first few hours could be due to the high sucrose content of fresh palm-wine which provides a good substrate for the growth of the bacterium and also a favourable almost neutral pH of the wine. The organisms in the palm-wine are alive and active (Ekundayo, 1969; Faparusi, et al., 1973; Rose, 1977; Swings and De Ley, 1977; Okafor 1977), and they metabolise the sugar present in the wine to produce alcohol and organic acid with a consequent decrease in pH of the palm-wine samples as shown in Table 1. With the decrease in pH and depletion in the substrate and growth substances, the growth of the bacterium becomes inhibited, hence the subsequent decrease in the number of the organism after 15hrs. of fermentation. The stage at which this bacterium reached its maximum density during palm-wine fermentation has not been reported (Swings and De Ley, 1977). Z. mobilis has good growth at higher alcohol concentration than is normally present in palm-wine (Fig. 2).The bacterium grew well in media with 2.5 – 10% alcohol. Normal palm-wine contains approximately 0.5% to 7. 1% ethanol depending on its stage of fermentation (Bassir, 1968; Van Pee and Swing, 1971; Okafor, 1978). Further increase in alcohol concentration beyond 10% resulted in poor growth of the bacterium. Swings and De Ley (1977) had shown that Zymomonas was alcohol tolerant. The growth of the organism in acidic and alcoholic conditions shows that the organism participates in the second stage of palm-wine fermentation (Collard and Levi, 1959; Bassir, 1962; Tortora et al., 1998; Ingraham and Ingraham, 2004). The pH of the fermenting wine changes from neutral to acidic (Table 1). The growth of the isolates at different pH values is as shown in Fig.3. Z. mobilis grows best at pH between pH 5 and 7. The pH of the palm-wine sample, 15h after collection (Table 1) which had the highest population (Fig. 1) also falls within this range. However, the optimum growth at pH 7.0 (Fig. 3) is in contrast to the growth of the organism in palm-wine sample after collection (Fig. 1) at 0hr when the pH for the Ogba sample was pH 7.0 (Table 1). Under natural condition (Ogba sample) the native microflora are alive and active and there is keen competition for the nutrients present in the wine. As the wine age some microorganisms dies off and others become more active which is the succession of microorganism in the palm-wine at different times. It can be inferred from this that, Z. mobilis is most active after 12h of collection of palm-wines. The reduction in pH during storage can be attributed to the utilization of the sugars present in the wine and the production of CO2 and other organic acids into the medium. The wide range of acidic pH suitable for the growth of Z. mobilis is not surprising since the bacterium has been isolated from acid wines like Cider with pH values of 4.0 (Swings and De Ley, 1977). Z. mobilis grew best at temperatures of 30 – 35°C (Fig. 4). Its growth is inhibited at higher temperatures. Most microorganisms except the thermophilic ones do not grow well at temperatures much higher than 35°C. Generally, statistical analysis of the data obtained in this study showed that, there is no significant difference in the growth of Z. mobilis isolated from the different locations under varying conditions of pH, alcohol and temperature. This similarity in growth properties of all the isolates is not surprising since they were obtained from the same natural substrate (palm-wine). The activity and hence the fermentative contribution of an organism can be determined by the volume of gas produced (Bassir, 1962; Talaro and Talaro, 1999; Nester et al., 2004). The result obtained on the fermentative ability of Z. mobilis is shown in Table 2. Z. mobilis when introduced alone to a sugar substrate produced more gas than when S. cerevisiae alone was introduced. This showed that Z. mobilis is a better fermenter of palm-wine than S. cerevisiae. This study also showed that a combination of Z. mobilis and S. cerevisiae did not ferment as much as Z. mobilis alone. This probably indicated an antagonism between the two organisms in terms of competition for essential nutrients in the medium. From the study, it is seen that Z. mobilis contributes to the fermentation of palm-wine. Z. mobilis is known to be beneficial to man and cattle. Preservation of palm-wine, through inactivity of the microorganisms especially Z. mobilis, should be done about 15hrs. after tapping, when the density of Z. mobilis is at its peak. The beneficial effect of Zymomonas mobilis to man will best be tapped at this stage. Acknowledgement: I am grateful to Dr. Y.S. Izuagbe for his guidance. REFERENCES
Copyright 2005 - Journal of Applied Sciences & Environmental Management The following images related to this document are available:Photo images[ja05005t2.jpg] [ja05005f4.jpg] [ja05005f3.jpg] [ja05005f2.jpg] [ja05005t1.jpg] [ja05005f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}