 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Applied Sciences & Environmental Management, Vol. 9, No. 3, 2005, pp. 81-86 Differential Response of Silkworm, Bombyx mori L. to Phytoecdysteroid Depending on the Time of Administration K. SASHINDRAN NAIR1, MIAO YUN-GEN2; S.NIRMAL KUMAR1 1Central Sericultural
Research and Training Institute, Manandavadi
Road, Mysore-
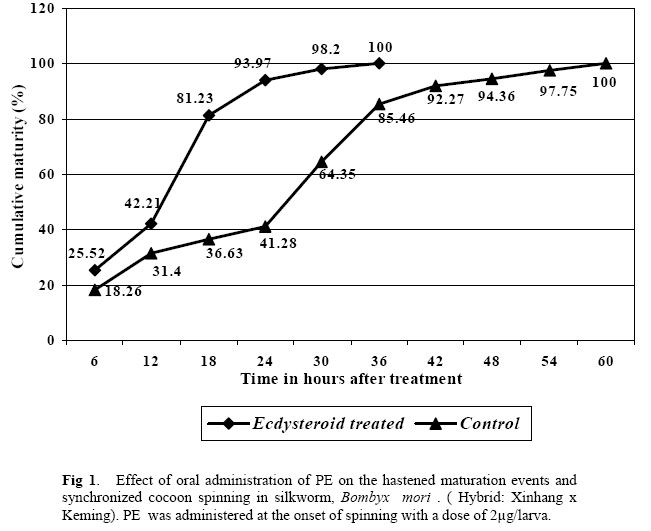
570 008, Karnataka, India. Email: nairjula@yahoo.com Code Number: ja05063 ABSTRACT: The differential response of silkworm, Bombyx mori L. to phytoecdysteroid when administered at different ages of 5th instar was studied on a popular Chinese silkworm hybrid, Xinhang x Keming reared on the ‘Hu Sang 197’ variety of mulberry leaves. Phytoecdysteroid, a chlolest -7 ene- 6- one- carbon skeleton (C27) from a plant source, Radyx achyranthes was administered to 5th instar silkworm per os at a rate of 2 µm per larva to different batches of silkworm at 48 h, 72 h and at the onset of cocoon spinning when a few larvae were ripe. The larval duration, larval mounting duration, cocoon characters and reeling traits were influenced by phytoecdysteroid. The intensity of influence was dependent on the time of application. The larvae treated at 48 h improved almost all economic traits significantly with a marginal increase in larval duration. In the larvae treated at 72 h, the economic traits were adversely affected but the larval duration was shortened by about 24 hours. In the larvae treated at the onset of cocoon spinning, the mounting duration was substantially reduced without much affect on the cocoon traits which would be a big benefit in commercial sericulture. The physiological significance of varied response of silkworm to phytoecdysteroid administration is discussed. @JASEM Ecdysone and juvenile hormone (JH) are the two major circulating hormones in insects which control majority of growth and developmental activities (Novak, 1975). Exogenous application of the analogues or mimics of these hormones could induce derangement in the metabolic activities and create disruptions in the normal insect development. More than the titre of these hormones, the balance between these two due to a well programmed interplay decides the pattern of physiological activities in insects at any given point of time. Plant-produced insect moulting hormones, termed phytoecdysteroids (PEs), function as plant defenses against insects by acting as either feeding deterrents or through developmental disruption (Schmelz et al., 2002). But the response of silkworm, Bombyx mori L. to minute quantities of these hormones or its analogues is beneficial. In China, various plant sources were identified which contained moderate to high amounts of PEs and used them in sericulture to manage the silkworm rearing during the last stage of larval development (Wong et al.,1979; Chow and Lu, 1980). The major objective of using PE in sericulture is to hasten the larval maturation events in the final larval instar and to synchronize the cocoon spinning process so that the larvae can be transferred early and together to the cocoon spinning device and the larvae would form cocoons almost simultaneously. This can save a lot of skilled labour which is otherwise required to pick up only the ripe worms in time and substantial amount of mulberry leaf. For such a result, PEs are usually administered at the last phase of the final instar when a handful of larvae are ripe. But the response of silkworms to administration of PEs at different ages of 5th instar is scant. This paper deals with the comparative response of silkworm to the application of PEs at 48 h, 72 h and at the onset of cocoon spinning. MATERIAL AND METHODS Experimental insect: Disease free eggs of bivoltine silkworm hybrid, Xinhang x Keming were used in this investigation. Silkworms were reared under standard recommended conditions at 26 ± 2 ° C temperature, 75 ± 5 % relative humidity under 12:12 (light:dark) photoperiod. ‘Hu Sang 197’ mulberry variety leaves, harvested from a frequently irrigated mulberry garden were fed to silkworms three times a day. After the third ecdysis, larvae were divided into four groups and reared under these same conditions. Administration of ecdysteroid: PE, a chlolest -7 ene- 6- one- carbon skeleton (C27) procured from Ningbo Biochemical Company, Ningbo, China was extracted from Radyx achyrancthes. Double distilled water was added to make a concentration of 20 µg PE per 1 ml solution. Silkworm larvae were allowed to feed on mulberry leaves sprayed with PE solution at the rate of 200 µg/10 ml water/100 g leaves/100 larvae. Such per os administration of PE was done to different batches of silkworm at 48 and 72 hours of 5th instar and at the onset of cocoon spinning. Untreated control was maintained in parallel to compare the results. Each treatment was replicated 5 times with 200 larvae per replication. Data collection: The duration of the 5th instar was calculated in the treated larvae and control. When the larvae started ripening, the rearing beds were examined every 6 hours, the ripe worms were collected, counted and transferred to mounting frames for cocoon building. Progressive maturation percentage was calculated and plotted in line graph. After cocooning, the cocoon weight, cocoon shell weight, shell percentage, filament length, denier, reelability and neatness were recorded/worked out. The data were subjected to statistical analysis employing ANOVA to ascertain the significance of the result. RESULTS AND DISCUSSION The data on the larval as well as larval mounting duration along with major cocoon characters are presented in Table 1. Data on reeling parameters are presented in Table 2. The cumulative maturation percentage of the PE treated larvae against the control has been plotted in Figure 1. Since the larvae treated at 48 and 72 hours of 5th instar did not show any difference from that of the control in cumulative maturation, only that of the larvae treated at the onset of cocoon spinning and the control has been plotted. The 5th instar larval duration and the duration to complete maturation process of all the larvae after a few larvae have been ripe were affected by the administration of PE. But this effect was largely dependent on the time of application. When administered to 48 h and 72 h old 5th instar larvae, the response was different from each others and the same was different from that of the larvae treated towards the end of the instar. In the first group in fact, the duration was extended by 6 hours compared to the control. But in the larvae treated at 72 hours, the 5th instar larval duration was shortened by 24 hours. In the third group, the larval duration was not affected as such since the application of PE was done only at the end of the instar but the duration of mounting was shortened substantially compared to control and that treated at 48 and 72 hours (Table 1). This is depicted clearly in Figure 1. The cumulative maturation percentage every 6 hours after the treatment shows that within 18 h of the treatment, about 81 % of the larvae matured whereas by the same time in the control only 37 % of the larvae matured. This sort of a reduction in the mounting duration in the PE administered silkworm reduces labour involvement in picking of the ripe worms and saves mulberry leaves. A similar result was reported by us by using PE extracted from Sesuvium portulacastrum on silkworm hybrids (Nair et al., 2002) and that from Caryophyllacea family of plants on pure silkworm breeds (Trivedy et al., 2003). This difference in the larval and mounting duration is because of a physiological role played by the exogenous ecdysteroid on the insect development system. The feeding larvae always contain a baseline level of ecdysone but reaches to pupation inducing peak before pupation (Sehnal, 1989). By giving an extra dose of plant based ecdysteroid at the critical time, the pupation inducing peak of ecdysteroid content in silkworm is advanced and thereby change the larval behaviour as such. Table 1: Effect of PE on the economic traits of silkworm, Bombyx mori (Hybrid: Xinhang x Keming) when administered at different ages of 5th instar. Values reported are the mean ± SD of 50 observations.
T48: Treatment at 48 h of 5th instar; T72: Treatment at 72 h
of 5th instar; Tsp: Treatment at onset of cocoon spinning The exogenous amount of PE also affects the larval weight, weight of the posterior silk gland, cocoon weight, shell weight and shell percentage as evident from Table 1. But the changes in these traits in response to PE treatment are dependent on the time of the treatment. The weight of mature larvae though did not change substantially, there was considerable difference. The larvae treated at 48 h gained in weight while those treated at 72 h and at the onset of spinning weighed less than control. Similar trend was also seen in the weight of posterior silk gland. The difference here was quite prominent. A gain of about 26 % was seen in T48 (treated at 48 h of 5th instar) while a reduction of about 13 % was seen in T72 (treated at 72 h). But in Tsp (treated at the onset of spinning) the reduction was marginal. Almost similar trend was obvious in cocoon characters as well. While there was considerable increase in cocoon weight, cocoon shell weight and shell percentage in T48 this was reversed in T72. In Tsp, the reduction was marginal and statistically non- significant. The changes seen in the reeling traits of cocoons formed out of treated and control silkworms also followed almost similar trend. The average and non breakable filament length had significant increase in the T48 whereas T72 showed significantly low values. The reelablity also showed a corresponding and complementary trend. The denier and neatness of the raw silk remained largely unaffected giving a bright and positive indication that the ecdysteroid treatments do not severely affect thickness and quality of the silk thread. Increase/decrease similar to the one seen in this study was reported by Chow and Lu (1980) when plant based ecdysteroid was administered to silkworm at early stages of 5th instar. The response of silkworm varied substantially depending on the time of treatment. The change in the reduction in the larval duration was inversely proportional to the cocoon characters as seen in the present study. These manifestations give an impression that PE administered early in 5th instar induces juvenoid like response in silkworm. Juvenoid treatment prolongs the larval period and increases the cocoon/silk weight in silkworm, B.mori. It was reported that silkworms are sensitive to exogenous ecdysteroid when administered at different hours (Sehnal and Akai, 1990). But such sensitivity and the related manifestations largely depend on the time of applications. Dai et al. (1985) indicated that ecdysone plays a significant role in nucleic acid metabolism and the related protein synthesis in silkworm. Although it induces growth and silk production, depending on the age of administration the intensity of manifestation will be different. According to Chow and Lu (1980) PE administered up to 48 h of 5th instar had induced an enhancement in the silk synthesis and the cocoon weight increased considerably. But the administration after 72 hours of 5th instar reduced the feeding period notably with an obvious reduction in the cocoon and cocoon shell weight. But the treatment at the onset of spinning brought about acceleration of maturation events and synchronization of spinning without a significant decrease in the quality of cocoons. The result of the present study agrees with these observations. Table 2: Effect of PE on the reeling traits of silkworm, Bombyx mori (Hybrid: Xinhang x Keming) when administered at different ages of 5th instar. Valuesreported are the mean ± SD of 50 observations.
T48: Treatment at 48 h of 5th instar; T72: Treatment at 72 h of 5th instar; Tsp: Treatment at onset pf cocoon spinning * Significant (P < 0.05); NS Non significant. The fact that effect of PE application is dependent on the time of application is fairly evident from the result of the 72 h treatment and that of the treatment made at the onset of spinning. The 72 h treatment is done obviously to save a silkworm crop from an imminent loss either due to an unforeseen shortage of mulberry leaves or disease outbreak, in the last phase of silkworm rearing. The treatment brings about a reduction of about 24 hours in the fifth instar period although it certainly reduces the economic traits as well. But the reduction in the cocoon crop is offset in this context because the crop would have been lost fully had the treatment been not made. In such event, the reduction in the cocoon traits is understandable since the final instar larvae do not get enough time to consume the leaves to the potential or as per its general requirement and thus the conversion to the silk protein is quantitatively affected though not qualitatively. The treatment done when a few larvae were ripe, presented a different picture. Here the thrust was more on the hastening of maturation process rather than attaining any improvement in the silk related economic traits. Hastened maturation and synchronized cocoon spinning save good amount of mulberry leaf and labour apart from enabling the farmer to market the produce in a single lot. Earlier, Shivakumar et al. (1995; 1996) reported such an effect on silkworm when extracts of plants such as Silene vulgaris, Silene inflata and Cassia tora were administered to silkworms. A physiological explanation for the varied effects of PE on silkworm could be in the following line: As proposed by Fukuda (1942), the increase in silk gland function during feeding period of the last larval instar is due to stimulation by ecdysteroid. For this, the origin of ecdysteroid need not necessarily be endogenous. It was also understood that the response of the silk glands to exogenous ecdysteroid depends on the developmental stage of the larvae (Akai and Kiuchi, 1988). Ecdysteroid represents a stimulator of silk gland. Feeding larvae always contain low level of ecdysteroid that may be indispensable for development (Sehnal, 1989). The dependence of silk production on ecdysteroid possibly reflects a general tissue requirement for such a low level of ecdysteroid concentration. A rise of ecdysteroid titre towards the end of fifth instar apparently terminates feeding and stimulates cocoon spinning in preparation to the metamorphosis. For proper course of these developmental events, it is significant that the ecdysteroid level is slightly elevated for 1-2 days before rising to the moult inducing height. The elevated titre of ecdysteroid apparently shifts silk glands to their regression phase when they reach maximum protein synthesis. The treatment, T72 just act this way and advances the moult inducing peak early. In Tsp, silk gland regression phase starts only after maximum silk gland growth is attained and hence the silk quantity remains almost unaffected. It can be surmised that PE is an effective bioactive compound which elicits responses in silkworm depending on the time of administration. Silk quantity can be increased by applying PE at about 48 h of 5th instar. The larval duration can be reduced at least by a day to save a crop from imminent loss by applying PE at about 72 h. The mounting duration can be reduced to about 24 h as against the regular 2-3 days and thereby labour involvement and leaf consumption. This study thus recognizes PE as one of the most important components in the silkworm rearing management. Acknowledgements: The author is extremely grateful to China Scholarship Council, Beijing, Govt. of China and the Ministry of Human Resource Development, Govt. of India, New Delhi, India for awarding the Chinese Government Scholarship under which the study was taken up. REFERENCES
Copyright 2005 - Journal of Applied Sciences & Environmental Management The following images related to this document are available:Photo images[ja05063f1.jpg] |
| |||||||||

{kind=link}