 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
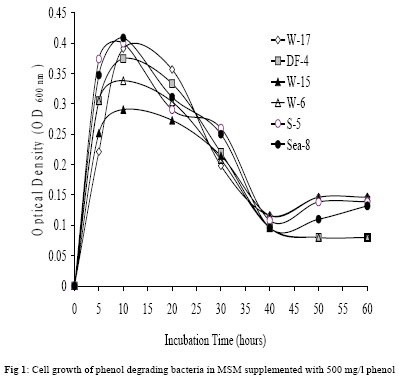
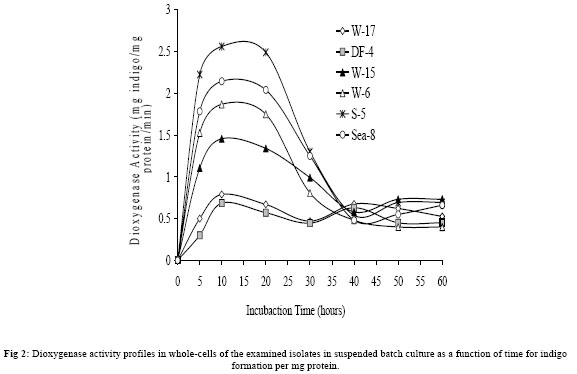
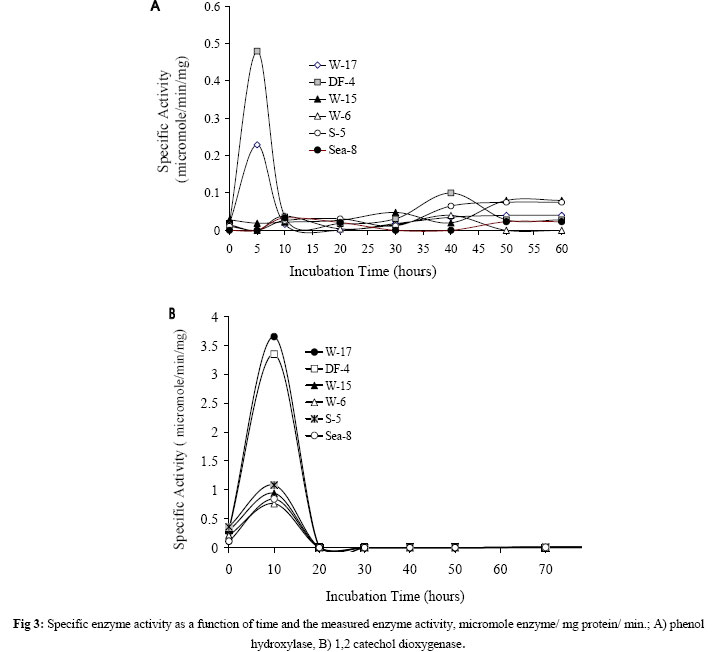
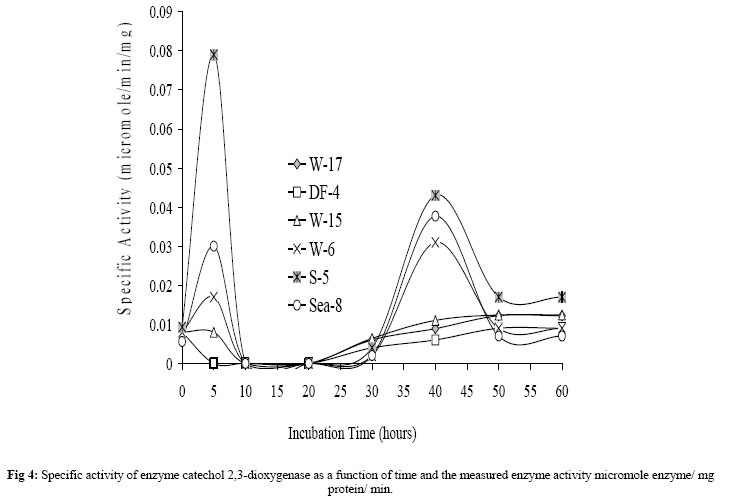
Journal of Applied Sciences & Environmental Management, Vol. 10, No. 3, September, 2006, pp. 75-81 Detection of meta- and ortho-cleavage dioxygenases in bacterial phenol-degraders *SAHAR ZAKI Environmental biotechnology Department, Genetic Engineering and Biotechnology Research Institute, MubarakCity for Scientific Research and Technology Applications, Post code 21934, New Burg-Elarab, Alexandria, Egypt. Code Number: ja06055 ABSTRACT: In the last five years, in our lab, several bacterial genera capable of degrading phenol as sole carbon source were isolated from different Egyptian ecosystems. Phenol mineralization using these isolates was ranged from 55% to 0.4%. In the present work, randomly chosen representative strains; W-17, DF4 (Acinetobacter), Sea-8 (Stenotrophomonas), W-6 (Klebsiella), S-5 (Bacillus) and W-15 (Ralstonia) were analyzed for the type of ring-cleavage(ortho or meta)dioxygenase physiologically induced during growth in thepresence of phenol as a sole carbon and energy source. The specific activities of the phenol-degrading enzymes phenol hydroxylase, catechol-1,2-dioxygenase and catechol-2,3-dioxygenase were investigated. In addition, whole-cell dioxygenases activities of the examined isolates were determined. Out of the results, only the Acinetobacter strains W-17 and DF-4 showed activity with the enzymes phenol hydroxylase and catechol-1,2-dioxygenase, which responsible for phenol degradation through ortho-cleavage pathway. In contrast, isolates S-5, Sea-8, W-6, W-15 and Pla-1 showed activity with the enzyme catalyzing the second step in the phenol degradation meta-cleavage pathway, catechol-2,3-dioxygenase. On the basis of our previous and present analysis, the investigated isolates are considered to have a good potential for application in remediation of phenol contaminated environments and industrial wastewater. @JASEM Phenol and phenolic compounds are of widespread use in many industries such as polymeric resin production and oil refining. As a result, these compounds are commonly encountered in industrial effluents and surface water. These pollutants are usually treated in activated sludge processes because many aerobic bacteria and fungi are able to use phenol as a source of carbon and energy (Rebhun and Galil 1988; Watanabe et al., 1996). Biodegradation of phenol, therefore, has long been the subject of numerous investigations (Ruiz-Rrdaz et al., 2001; Chang et al., 1998; Fava et al., 1995; Abd-El-Haleem et al., 2003; Dean-Ross, 1989; Solomon et al., 1994; Ahmed et al., 1995; Alleman et al., 1995; Collins and daugulis, 1997; Fulthorpe and Allen, 1995; Lin et al., 1990; Morris and Lester, 1994; Ryu et al., 2000; Wang et al., 1996). A typical pathway for metabolizing an aromatic compound like phenol is to dihydroxylate the benzene ring to form a catechol derivative and then to open the ring through ortho or meta oxidation. Catechol is either oxidized in a reaction catalyzed by catechol-1,2-oxygenase which is described as an ortho pathway, or is oxidized in reaction catalyzed by 2,3-dioxygenase the meta pathway to 2-hydroxymuconic semialdehyde. The final products of both the pathways are molecules that can enter the tricarboxylic acid cycle(Powlowski and Shingler 1994; Harayama et al. 1992). Previously in our lab twelve bacterial phenol-degraders which could grow aerobically and assimilate 500 mg/l phenol for a period of 15 h were isolated and Molecular characterized by Abd-El-Haleem et al. (2002 and 2003). Their comparative sequence analysis of the 16S ribosomal DNA, classified them to different affiliated groups. Four isolates of them W-17, DF-4, Sea-9 and Sea-12 were closely related to the genus of Acinetobacter. Two strains Sea-8 and Sea-11 were belonging to Stenotrophomonas maltophilia. Strains DM-5 and W-6 were affiliated to Klebsiella. Isolates S-5 and W-15 were closely related to Bacillus and Ralstonia respectively. Some of these isolates have been immobilized into organic and inorganic carriers and has shown good capacity for long-term degradation of high phenol concentration (Abd-El-Haleem et al. 2003; Beshy et al. 2003). In the present study randomly selected representative strains; W-17, DF4 (Acinetobacter), Sea-8 (Stenotrophomonas), W-6 (Klebsiella), S-5 (Bacillus), W-15 (Ralstonia) and Pla-1 (Microbacterium) were analyzed for the type of ring-cleavagedioxygenase physiologically induced during growth in thepresence of phenol as a sole carbon and energy source. The specific activities of the phenol-degrading enzymes were investigated. MATERIALS AND METHODS Materials: Unless specified otherwise, all reagents were of the highest purity available. Bacterial strains:Isolation and molecular characterization of the all bacterial strains used in this study described previously by Abd-El-Haleem et al. (2002). Medium and culture conditions: To achieve enough amounts of cells, isolates were grown in LB-medium, which contained per liter: 5 g yeast extract, 10 g casein peptone, 10 g NaCl, while the pH was adjusted to 7.0 before autoclaving. Subsequently, 10 % inocula from LB cultures were transferred to 50 ml freshly prepared minimal salts medium {(MSM) containing (per liter) 2.75 g of K2HPO4, 2.25 g of KH2PO4, 1.0 g of (NH4)2SO4, 0.2 g of MgCl2.6H2O, 0.1 g of NaCl, 0.02 g of FeCl3.6H2O, and 0.01 g of CaCl2 (pH 6.8 to 7.0)} supplemented with 5 mM phenol as sole carbon and energy source. All growth experiments were run in 250 ml Erlenmeyer flasks containing 50 ml of medium at 30 °C on a rotary shaker at 200 rpm. Enzyme assays:All enzyme assays were routinely performed at 30°C using a Beckman DU7500 spectrophotometer with integral chart recorder and temperature control. All enzyme rates are the average of at least three determinations. The enzyme activity was defined as the initial rate of indigo formation or enzyme specific activity normalized to the protein content of the sample. Determination of normalized dioxygenases activity in whole cells: The optical density of a 1 ml sample of the culture was determined; then the sample was immediately transferred to a microcentrifuge tube. The cells were harvested and washed with phosphate buffer solution (pH=7.2) by centrifugation (1 min at 14 000 rpm at 4°C). The enzyme reaction was initiated by addition of indole (Jenkins and Dalton, 1985). Five microliters of 100 mM indole in N,N-dimethlylformamide was added to the cells and the formation of indigo was monitored spectrophotometrically at 600 nm over the reaction time against a control (resuspended cells without indole). The initial rate of indigo formation was determined by plotting the increase in indigo absorbance as a function of time. Phenol hydroxylase:The oxidation of NADH in the presence of phenol by hydrogen peroxide-treated cell extracts was monitored at 340 nm. Reaction mixtures (3 ml) containing 50 mm KH2PO4: K2HPO4 buffer pH 7.2, 1.0 mmol NADH, and 100 nmol phenol were equilibrated at 55°C before the addition of the cell extract (100 ml). One unit of activity was defined as the amount of enzyme catalyzing the oxidation of 1 mmol NADH min (Gibson et al. 1990). Catechol-2,3-dioxygenase:Reaction mixtures (3 ml) in 50 mm KH2PO4:K2HPO4 buffer pH 7.2 containing 1 mmol catechol were equilibrated at 55°C before adding the cell extract (100 ml). The increase in absorbance at 375 nm caused by the formations of the reaction product 2-hydroxymuconic semialdehyde was monitored (Masai, et al. 1995) One unit of enzyme activity was defined as the amount of enzyme catalyzing the production of 1 mmol 2-hydroxymuconic semialdehyde per minute at 55°C. Catechol-1,2-oxygenase:Reaction conditions were identical to those of the catechol-2,3-dioxygenase assay except the formation of cis, cis-muconic acid which was monitored at 260 nm (Ornston, et al. 1966) Protein concentration:Protein concentrations were measured by the Bradford method (Bradford, 1976) using the Bio-Rad protein assay reagent. Bovine serum albumin (Sigma, Fraction V) was used as a standard. RESULTS AND DISCUSSION A representative growth profile is shown in Fig 1, periodic monitoring of cell density revealed differences in growth characteristics between the cultures. All isolates reached their maximum growth after about ten hours of incubation. Thereafter, the growth rates of all isolates were approximately constant till 30 hours of incubation, then decreased dramatically until the end of the experiments (60 hours of incubation).The conversion of indole to indigo was used as an indicator of dioxygenase activity in the whole-cells. Therefore, all isolates were assayed for indigo production. As shown in Fig 2, whole-cell dioxygenase activity profiles were correlated with growth of the isolates. Dioxygenase activity was observed to vary from low to high between 0.3 (isolate DF-4) and 2.22 (isolate S-5) mg indigo/mg protein/min during the log growth phase (after incubation time for ~ 5 hours). During the stationary growth phase (from ~ 10-20 hours incubation), the enzyme activity increased to between 0.68 (DF-4) and 2.56 (S-5) mg indigo/mg protein/min. Hugouvieux-Cotte-Pattat et al. (1990) reported a similar growth-phase-dependent expression of the catabolic genes by P. putida. Phenol-degrading aerobic bacteria are able to convert phenolinto nontoxic intermediates of the tricarboxylic acid cyclevia an ortho or meta pathway (Powlowski and Shingler, 1994). The first step of both routesis the monohydroxylation of the ortho position of the aromaticring (Harayama et al. 1992). The enzyme responsible for the hydroxylations at the positionsortho is the monooxygenasephenol hydroxylase. It is also known that the efficiency of a certain catabolic pathway often depends on the properties of the involved key enzyme(s).Therefore, the specific activities of phenol hydroxylase and catechol 1,2-dioxygenase in cell-free extracts obtained by ultrasonication from investigated isolates were examined. As shown in Fig 3 A, B, the enzymes phenol hydroxylase and catechol 1,2-dioxygenase of exhibited certain similarities and differences between examined microorganisms. The data showed also that the highest activities of both enzymes were in the Acinetobacter isolates DF-4 and W-17, while no phenol hydroxylase activates were detectedunder similar conditions in the rest of the examined isolates. These results suggest phenol metabolism throughout occurred via ortho fission of catechol in the Acinetobacter isolates. In addition, isolate W-17 that presented the highest phenol mineralization rate (55%) as reported previously by Abd-El-Haleem et al. (2002), presented higher activities of both enzymes phenol hydroxylase and catechol 1,2-dioxygenase, respectively. In contrast to the above finding, a large amount of meta-cleavage dioxygenase (catechol2,3-dioxygenase) activity was detected in the cells of isolates S-5 (Bacillus), Sea-8 (Stenotrophomonas), W-6 (Klebsiella) and W-15 (Ralstonia) respectively in order of decreasing activity; while no meta-cleavage dioxygenase activity was detectedunder similar conditions in the Acinetobacter isolates W-17 and DF-4 (Figure 4).Previously, extradiol catechol 2,3 dioxygenases have been purified from a variety of organisms comprising Pseudomonas, Alcaligenes and Bacillus (Ornston and Stanier 1966; Kataeva and Golovleva 1990; Kang et al. 1998; Chen et al. 1998). On the basis of our previous and present results, the investigated isolates are considered to have good prospects for its application for microbial detoxification of phenol pollutes industrial wastewater. A general comparison of the major pathways for catabolism of aromatic compounds in bacteria has revealed that the initial conversion steps are carried out by different enzymes but that the compounds are transformed into a limited number of central intermediates, such as protocatechuate and (substituted) catechols (Van Der Meer et al. 1992;Chaudhry and Chapalamadugu 1991). These dihydroxylated intermediates are channeled into the ortho cleavage pathway or meta cleavage pathway (Powlowski and Shingler 1994; Harayama et al. 1992). Both types of pathways lead to intermediates of central metabolic routes, such as the tricarboxylic acid cycle. Finally, it may possible to conclude that some bacteria employ more than one pathway to degrade hydrocarbons. These can easily allow the formation of new mixed metabolic pathways and might explain why bacterial strains capable of growing on contaminants emerge so quickly. REFERENCES
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}