 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
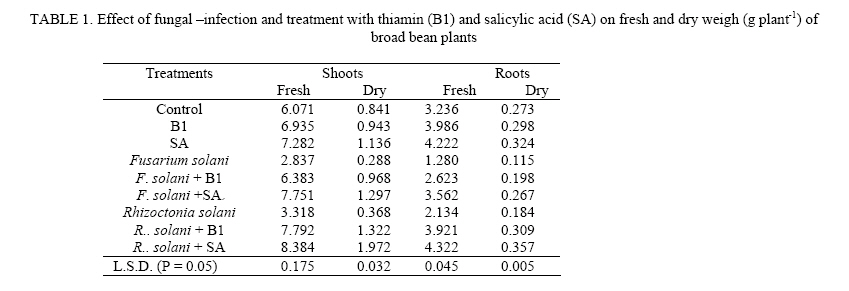
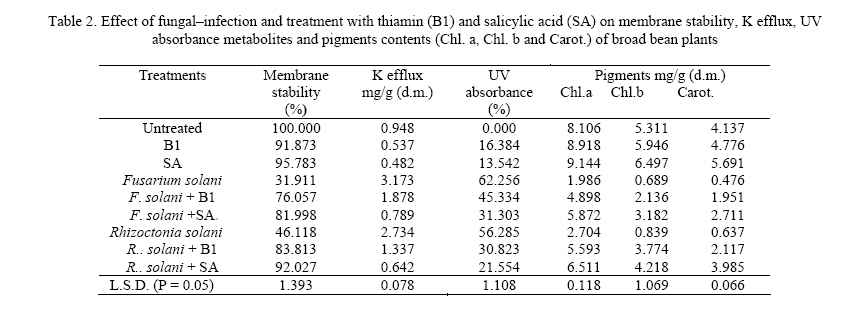
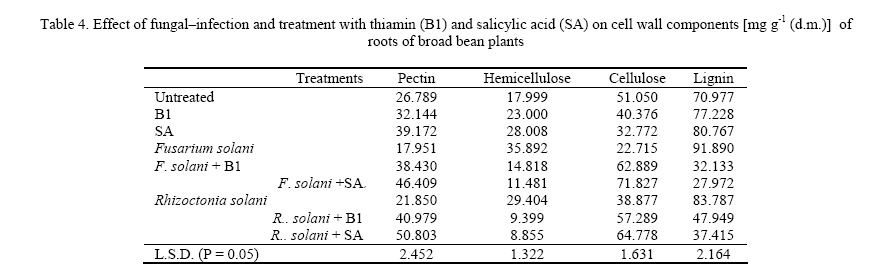
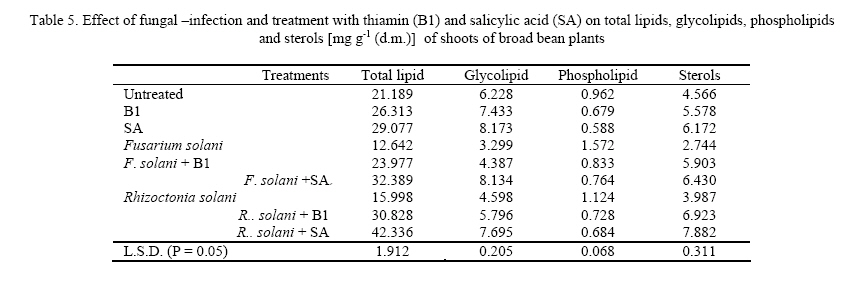
Journal of Applied Sciences and Environmental Management, Vol. 11, No. 4, 2007, pp. 125-131 Thiamin and salicylic acid as biological alternatives for controlling broad bean rot disease 1AL- HAKIMI, A M A; 1*SAEED M.S. ALGHALIBI 1Department of Biology, Faculty of Science, Taiz University,Taiz, Yemen Code Number: ja07108 ABSTRACT: The interactive effects of fungi (Fusarium solani and Rhizoctonia solani) infection and thiamin or salicylic acid on growth rate, membrane stability, K+ efflux, UV-absorbing metabolites, photosynthetic pigments, cell wall components and lipid fractions of broad bean plants (30-day-old) were studied. Fungal infection induced a reduction in growth rate, membrane stability and content of photosynthetic pigments. Foliar application of thiamin or salicylic acid increased growth rate, membrane stability and content of photosynthetic pigments. The K+ efflux and the leakage UV-absorbing metabolites were stimulated with fungal infection. However, thiamin and salicylic acid treatment partially retarded the stimulatory effect on leakage of K+ and UV-absorbing metabolites of fungal infected plants. Fungal infection produces a reduction in the content of pectin and cellulose, total lipid, glycolipids and sterols fraction of shoots and roots and phospholipids of roots. On the other hand, the contents of hemicellulose and lignin of shoots and roots and phospholipids of shoots were stimulated by fungal infection. Soaking seeds in thiamin or salicylic acid counteracts partially or completely the adverse effect of fungal infection on pectin and cellulose composition, total lipid, glycolipids and sterols of either shoots or roots. On the other side, thiamin or salicylic acid treatments retarded the phospholipids accumulation in shoots of infected plants, and in roots the phospholipids accumulation was partially or completely alleviated. The content of hemicellulose and lignin of shoots and roots were antagonistically lowered by the application of thiamin or salicylic acid. With the increase concern of conserving natural resources and reducing air, and water pollution, natural or biological control of plant diseases had increased emphasis (Hamada and Hashem, 2003). Biological control of plant diseases is show, gives few quick profits, but can be long lasting, inexpensive and harmless to life (Dhingra and Sinclair, 1994). Responses mediated by thiamin and salicylic acid suggested that these plant-derived substances have important physiological roles and great potential as elicitors and mediators of resistance signal transduction. The two compounds induce unique type of resistance when topically applied, and they affect a variety of processes in plants, including environmental acclimatization and defense against pathogens (Delancy et al. 1994; Hashem and Hamada, 2002). The main effect of these compounds against plant pathogens is that they induce the resistance of plants against pathogens and stimulate plant growths which recover any disturbance produced from the plant pathogens (Nickell, 1983; Vidhyasekaran, 1990). The recent studies indicated that, treating plants with some vitamins resulted in pathogen resistance. Foliar application of riboflavin (B2) effectively controlled several diseases of tobacco (Dong et al., 1995), and reduced the powdery mildew of strawberry plants in combination with methionine (Wang and Tzeng, 1998). Enkerli et al. (1993) reported that, salicylic acid is the only plant-derived compound induced defense response known as systematic acquired resistance (SAR). Thus, in this work it seemed necessary to study the effects of fungal infection on growth rate and some metabolic processes and the role of thiamin or salicylic acid in amelioration of these adverse effects of fungal infection. MATERIALS AND METHODS Fusarium solani and Rhizoctonia solani were kindly supplied by Agriculture Research Center. Ministry of Agriculture, Giza, Egypt. Fusarium solani and Rhizoctonia solani were grown first in slants containing potato dextrose agar medium. A spore suspension was prepared in sterile water from 5 to 6 days old culture of Fusarium solani and Rhizoctonia solani. Potato dextrose agar plates were then inoculated with one ml of spore suspension and incubated at 27° C for 14 days. At the end of the incubation periods four discs (0.5 mm in diameter) were taken from 14 days old culture and used to inoculate 500 ml bottles containing soaked sorghum grains and sand with ratio 4: 1 and incubated at 27 ° C for three weeks and used as fungal inoculums. Fungal inoculums were added with cultivation in pots with ratio 3- 5%. Broad bean seeds were soaked either in 0.3 mM thiamin (B1) or in 0.6 mM salicylic acid (SA) before sowing for 6 hours. Five soaked seeds were sown in pots infested with the pathogens. The soaked seeds were also in anther groups of pots without addition of pathogens. Non-treated seeds were planted in non-infested pots to serve as a control. All treatments were in 3 replicates and arranged in complete randomized design. At the end of the experimental period (30 days) fresh shoots and roots were then dried in an aerated oven at 70° C until constant weight. Cell membrane stability was determined according to the method of Prmachandra et al. (1992). Leaf discs (number of 10, each 1 cm diameter) was rinsed three timeswith deionized water and placed in 30 cm3 deionized distilled water for 24 h at 10° C. The electrical conductivity (conductimeter, YSI Model 35 Yellow Springs, OH, USA) of the bathing solution was measured at 25 0C. Following the measurements, leaf discs were autoclaved for 15 min, cooled to 25° C, and the electrical conductivity of the bathing solution was measured for the second time. The degree of injury was calculated according to the equation: Percentage injury (%) = 1- (1-T1/T2) / (1-C1/C2) ×100 Where T1 and T2 are the first (before autoclaving) and second (after autoclaving) conductivity measurements of the treatment, respectively, C1 and C2 are the first and second conductivity measurements of the control. The flame photometric method (Williams and Twine, 1960) using Carl Zeiss flame photometer was used for the determination of potassium. The leakage of UV absorbing metabolites according to Navari-Izzo et al. (1989) was also used as a criterion for evaluating cell membrane integrity of leaf discs. The data were then expressed as A280 (10 cm3)/g fresh matter after 24 h leakage in distilled water. The content of chlorophyll a, b and carotenoids was determined by the method of Metzner et al. (1965). Cell wall fractionation was conducted essentially according to Dever et al. (1968) and Galbraith and Shields (1981). Tissue powder samples were extracted twice in distilled water, twice in 80% ethanol to remove soluble sugars metabolites. The residue was then extracted in 2 ml 0.5 N NaOH for starch, 0.5% ammonium oxalate-oxalic acid (90 ºC for 24 h) for pectin, 17.5% NaOH for hemicellulose and in 72% H2SO4 (with 15 min autoclaving) for cellulose extraction. After that, the remaining precipitate was ascribed to the lignin fraction according to Dever et al. (1968). Content of wall polysaccharides were determined by the anthorne sulfuric acid reagent using glucose as standard (Fales, 1951). Lipids were extracted three times for dried plant organs (each lasted for 24 h) with chloroform/methanol (2:1, v/v) at room temperature according to Navari-Izzo et al. (1989). Lipids (100 µg) were chromatographed on silica gel plates (silica gel G/60) using n-hexine-diethylether-glacial acetic acid (70: 30: 1) and the spots were visualized by I2 vapor. According to Navari-Izzo et al. (1989) the total polar lipids (TPL) were located at the origin of the chromatogram. These were scratched and redissolved in chloroform/methanol (2:1). Glycolipids were estimated as their monosaccharide content by the anthrone sulfuric acid reagent. Phospholipids were determined in lipid extracts according to Johnson (1971). Lipid phosphorus was estimated by the molybdate blue colour (Woods and Mellon 1941). Phosphorus content was taken as an index for phospholipid contents. Total sterols were estimated according to the method described by Cook (1958). Data obtained in the above mentioned tests were analyzed statistically, and the mean were compared using L.S.D. test (Mead and Curnow, 1983). RESULTS AND DISCUSSION The fresh and dry weights of both shoots and roots (Table 1) were significantly reduced by fungal infection. These results were confirmed by finding of many researches (Badr-El-Din and Moawad, 1998; Hashem and Hamada, 2002; Hamada and Hashem, 2003; Metwali et al., 2005). Soaking of broad bean seeds in thiamin or salicylic acid did not only alleviate the inhibitory effects of fungal infection but also were of stimulatory effects, where the fresh and dry matter seed in shoots and roots showed a marked increase (Table 1). The results proved our previous finding (Enyedi et al., 1992; Spletzer and Enyedi, 1999; Hamada and Hashem, 2003). Membrane stability clearly showed a highly significant decrease with fungal infection (Table 2). A similar reduction in membrane stability induced by fungal infection stress has also been observed by Rhodes (1987), Mednetsev et al. (1988) and Hashem and Hamada (2002). Seed soaking in thiamin or salicylic acid was generally effective in antagonizing partially the inhibitory effects of fungal infection stress on the membrane stability Thiamin is an antioxidant (Frederikse et al., 1999) and other antioxidants induced disease resistance in plants (Norris and April, 1991). Also, salicylic acid played an important role in plant defense by the development of a systemic acquired resistance against pathogens (Ryals et al., 1994) and by increasing antioxidant enzymes (Janda et al., 1999). With respect to the K+ efflux and the leakage of UV-absorbing metabolites, the results presented in table 2 revealed that the K+ efflux and the leakage of UV-absorbing metabolites were stimulated with fungal infection. Such leakage of ionic solutes and potassium efflux (Hashem and Hamada, 2002) as well as leakage of cellular metabolites (Navari-Izzo et al., 1993; Hamada and Hashem, 2003) are frequently used to asses membrane integrity. In this respect, Nemec (1995) concluded that fungal phytotoxins enhanced membrane permeability. The applied thiamin or salicylic acid was generally effective in antagonizing partially the stimulatory effects of fungal infection stress on the leakage of K+ efflux and UV-absorbing metabolites. These results are in accordance with results obtained by (Hamada and Hashem, 2003). The data herein obtained Table 2 clearly demonstrated that the fungal infection had inhibitory effects on pigment fractions (chlorophyll a, b and carotenoids) biosynthesis in broad bean leaves. These results are agreement with Albrecht et al. (1998) who showed that the F. solani toxin dihydrofusarubin caused degradation of leaf pigments close to the veins of tobacco leaves treated with the toxin in light, thus indicating that dihydrofusarubin acts a non-host specific toxin. Also, they demonstrated that dihydrofusarubin interacted with the electron transport chain of illuminated spinach chloroplast, resulting in the formation of super oxide radicals-generated by the photosystem I. The beneficial effects of the applied thiamin or salicylic acid in mitigative partially or completely the adverse effects of fungal stress on photosynthetic pigments were clearly exhibited by the test plant. The mitigation of the harmful effects of fungal stress on plant growth due to thiamin or salicylic acid treatments could be directly attributed to the role of the applied thiamin or salicylic acid in enhancing the photosynthetic activities and chlorophyll biosynthesis. These results are agreement with Neubauer and Yamamota (1992), Chloudhury et al. (1993), Frederikse et al. (1999) and Hamada and Hashem, (2003) who attributed positive effects of thiamin and salicylic acid for stabilizing and protecting the photosynthetic pigments and the photosynthetic apparatus from being oxidized. Cell wall structure and properties determine, to a large extent, the magnitude of cell division and elongation, and hence plant growth. Therefore, cell wall components of the studied plants were estimated to evaluate how far they could have been affected by fungal infection stress. The accumulation of pectin and cellulose fractions in broad bean shoots and roots were significantly lowered by fungal infection stress, whereas hemicellulose and lignin of either shoots or roots were stimulated (Tables 3, 4). Treatment with thiamin or salicylic acid stimulated the accumulation of pectin and cellulose in shoots and roots of infected plants. Furthermore, the data clearly demonstrated the capability of thiamin or salicylic acid treatment in retarding the stimulatory role of fungal infection on the production of hemicellulose and lignin in shoots and roots of the infected plants. Plants have generally evolved complex mechanism including both passive (preformed) and active (inducible) defense responses for protection against pathogenic agents. The active defense responses include rapid modification of existing cell wall material, and deposition of new cell wall material including lignins, callose, phenolics and hydroxyproline-rich glycoproteins (Darvill and Albersheim, 1984; Lamb et al., 1989; Cramer and Radin, 1990; Bradley et al., 1992; Hamada et al., 2001). The results also clearly indicate the capability of thiamin or salicylic acid in mitigating partially or completely the depressive effect of fungal infection on the cell wall component of the test plants. The mitigative effect of seed imbedded in thiamin or salicylic acid on the stimulated pectin and cellulose accumulation of the test infected plants and inhibited lignin production the infected plants may be one aspect of the role of thiamin or salicylic acid which should be considerable helpful in defense and concomitant fortification of the plant via the cell wall. Lipid and lipid fractions of the studied plants exhibited variable changes in response to the fungal infection and their interaction with the applied thiamin or salicylic acid (Tables 5, 6). Total lipid contents of shoots and roots were significantly decreased with fungal infection. Fungal infections significantly decreased glycolipid and sterols fraction of all organs of the test plants. The phospholipid fraction was significantly increased in shoots, whereas it decreased in roots of infected plants. Phospholipids play an important role in membrane permeability, mitochondrial electron transport (Green and Tzagoloff, 1966) and subsequently any reduction in phospholipids will affect a variety of organelle functions. Foliar application of thiamin or salicylic acid not only relieved the inhibitory effects of fungal infection but also had of stimulatory effects, where the total lipids, glycolipids and sterols in shoots and roots showed a manifest increase. On the contrary, thiamin or salicylic acid treatments retarded the phospholipids accumulation in shoots of infected plants, while in case of roots the phospholipids accumulation was partially or completely alleviated. Membrane lipids form a physical barrier to the movement of the water soluble components of the cells. The remarkable changes in lipid and lipids fractions of the test plants may also be due to the fungal infection which damages the cell plasma membranes. Similar decrease or increase in lipid contents by stress imposition were also recorded in plants (Dakhma et al., 1995; Quartacci et al., 1997; Hamada et al., 2001; Al-Hakimi 2003, 2005). The decrease in phospholipids may be due either to reduction in the power of hydrophobic proteins to bind phospholipids (Heber and Sanatarius, 1964) or due to lipid peroxidation on enhanced lecithinase (Harrison and Trevelyan, 1963). In this respect, Hamada et al. (2001) reported a general decrease in phospholipid and glycolipid contents with nematode infection. Sterols also play essential functions in the lipid core of cell membrane, and they are also biogenetic precursors of numerous secondary metabolites including plant steroid hormones (Geuns, 1978). It has been shown that sterols could influence the structural and functional properties of biological membranes (Horvath et al., 1981). REFERENCES
Copyright 2007 - Journal of Applied Sciences & Environmental Management The following images related to this document are available:Photo images[ja07108t5.jpg] [ja07108t6.jpg] [ja07108t1.jpg] [ja07108t4.jpg] [ja07108t2.jpg] [ja07108t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}