 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
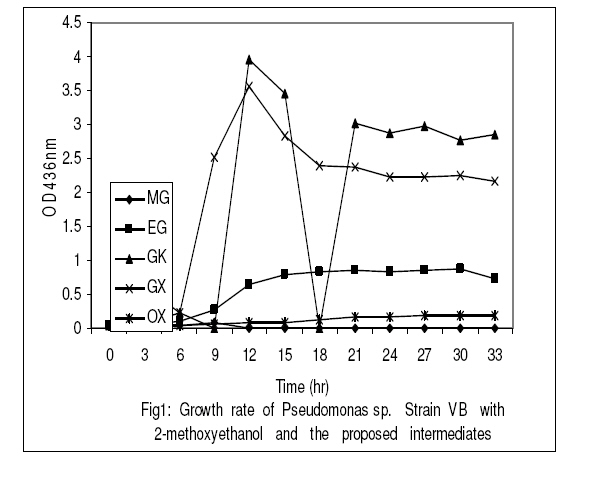
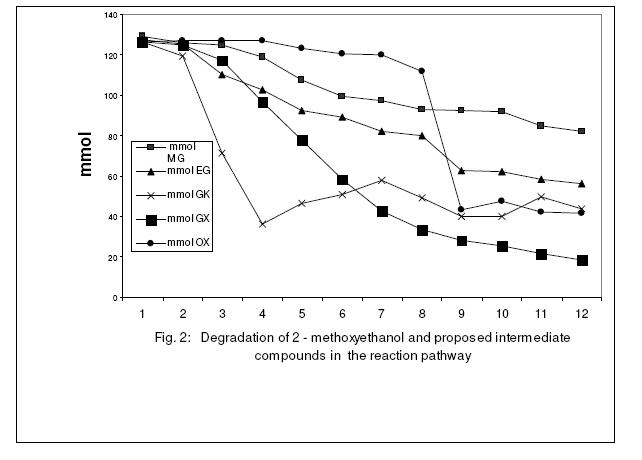
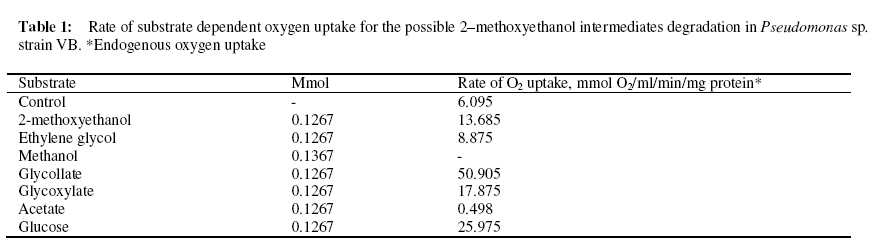
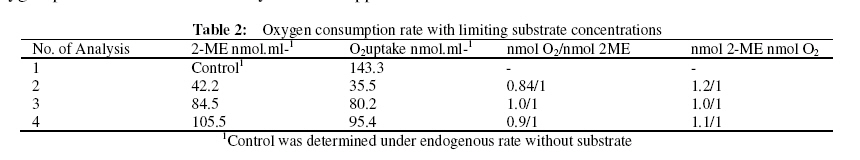
Journal of Applied Science & Environmental Management Vol. 15 No.1, March 2011, pp. 51 -55 Biodegradation of 2 -methoxyethanol by a new bacterium isolate Pseudomonas sp. Strain VB under aerobic conditions 1Ekhaise, F O; 2Meyer, O 1Department of Microbiology, Faculty of Life Sciences, University of Benin, Benin City, Nigeria, Code Number: ja11011 ABSTRACT Microbial biodegradation of 2-methoxyethanol also known as Methyl glycol (MG) under anaerobic conditions has received much attention during the past decade. However, not much is known about the aerobic degradation of 2-methoxyethanol. Samples from various environmental niches were enriched to isolate and determine bacterial isolates capable of utilizing 2-methoxyethanol as a sole source of carbon and energy under aerobic conditions. A 2-methoxyethanol degrading bacterium was isolated from anaerobic sludge of a municipal sewage from a treatment plant in Bayreuth, Germany, by selective enrichment techniques. The isolate was designated strain VB after it was shown by the 16S rRNA phylogenetic sequence analysis as belonging to the genus Pseudomonas. Under aerobic conditions Pseudomonas sp. strain VB was capable of mineralizing 2-methoxyethanol and its intermediary metabolites. Stoichiometrically, the strain utilized one mole of oxygen per one mole of 2-methoxyethanol instead of four mole oxygen per one mole of 2-methoxythanol for the total oxidative metabolism. 2-methoxyethanol (C3H802) (methyl cellosolve) is a colourless substance that is highly soluble in water and acetone, with a molecular weight of 76.10g/mol and density of 0.96g/1. It is used in nail polishes and wood staining techniques. It functions as a good extracting agent for a number of compounds of PCE, BTEX-aromatics and toluene (Brau-Stromeyer and Meyer, 1995). 2-methoxyethanol is poisonous when ingested orally and has a lethal concentration of 1500ppm for mice in air (Chemie, 1965; Merck, 1989). It impairs fertility and causes harm to the inborn child when pregnant mothers are exposed to it. The critical role of microorganisms in degradation of organic pollutants is well known. Although the microbial mixed cultural degradation of 2-methoxyethanol under anaerobic conditions has in the past decades been extensively investigated (Tanaka et al., 1986; Tanaka and Pfennig, 1988). Information on the microbial degradation of 2-methoxtethanol by a single bacterium isolate under aerobic conditions remains rarely investigated (Ekhaise, 2002). In the present study, search for a single bacterium that can degrade 2-methoxyethanol aerobically will be made with a view to elucidate the microbial degradation pathway of the compound. MATERIALS AND METHODSIsolation and Growth Criteria: The mineral medium (Meyer and Schlegel, 1983) used in this study consisted of Na2HPO4x 7H20 9.0g, KH2PO4 1.5g, NH4Cl 1.5g, MgSO4 x 7H2O 0.2g, CaCl2 x 2H2O 20.0mg, Iron (III) ammonium citrate 1.2mg, and trace element solution TS2 1.0ml per 1 litre of deionized water. The pH value of the medium was adjusted with 10 N NaOH to 7.2 before autoclaving at 121°C for 20min. Solid medium was prepared with 1.5% (W/V) Agar and 0.5% (w/v) each of nutrient broth and yeast extract. Enrichment and Isolation of 2-methoxyethanol utilizing bacterium: In order to isolate a bacterium strain capable of utilizing 2-methyoxyethanol as the sole source of carbon and energy, microbial consortium from waste – waters were used for direct inoculation of 100ml mineral medium in 300ml Erlenmeyer flask. The medium was supplemented with 2-methoxyethanol (126.7mmol). Cultures were grown aerobically on a rotatory shaker 200 rev/min at 300C for 24hrs. Culture showing growth was transferred to fresh medium, after 3 – 5 successive transfers, the enrichment cultures were plated onto solid medium. After 2 – 3 days of incubation, the plates were examined for growth. Single colonies were picked and streaked for purity. This step was repeated until homogenous colonies were obtained. These colonies were inoculated into the liquid medium supplemented with 2-methoxyethanol to ensure their biodegrdative ability. The morphological physiological and biochemical characterization of the isolate was examined according to the methods of Gerhardt et al. (1994). The bacterium strain was identified by the 16S rRNA sequencing and analysis of the bacterial isolate as described by Rainey et al. (1996). The sequencing and analysis of the 16S rRNA of the bacteria strain was determined at the Deutsche Sammlung von Mikroorganismen and Zellkulturen (DSMZ) Braunschweig, Germany. Utilization of 2-methoxyehanol as a sole source of carbon and energy: The utilization and microbial biodegrading of 2-methoxyethanol was studies with Pseudomonas sp strain VB in mineral medium with the initial concentration of 126.7mmol of the compound per 100ml. Exponentially growing cells from the mineral medium supplemented with 2-methoxyethanol were used as inoculums. The experiment was performed in 10L Fermetor with 8L mineral medium at a gas flow rate of 5.5L/min (The laboratory house-built compressed air was used as the source of oxygen). Cultures were grown aerobically with a rotatory shaker at 200 rev/min and a growth temperature of 30° C. Ten milliliters of the culture medium were taken periodically and the growth rate was monitored turbidometrically at optical density (OD) of 436nm with a Spectrophotometer (LKB Biochrom Ultrospec 4050 Cambridge, England). The withdrawn potions of the samples were centrifuged (10,000xg for 4min) in microfuge to remove cells and the supernatant were analyzed for the mineralization of 2–methoxyethanol by gas chromatography. Oxygen consumption assay with resulting cells: Cells were harvested by centrifugation for 10min at 10,000xg and 4° C from overnight shown culture with 2–methoxyethanol, and washed twice with 50mM phosphate bugger (pH 7.2) washed cells were suspended in the same buffer. The determination of oxygen uptake rates was performed polarographically at room temperature by using a Clark -Type electrode Model 10 Ranke Brothers, Birmingham, Great Britain) connected to a printer (Packard Model 13, Kipp and Zonen, Netherland). The assay was performed with the washed resting cells suspended in 50mM phosphate buffer (pH 7.2). Oxygen consumption or uptake was measured in 3ml of 40mM phosphate buffer (pH 7.2) at room temperature with a magnetic stirrer with Clark – Type electrode. The buffer was saturated with compressed air from in-built laboratory gas flow for at least 5min before finally the reaction chamber was closed without air bubble. The reaction was initiated with the addition of 100 µ l of an intact cells suspension and 0.126mmol concentration of the potential metabolites. The oxygen uptake was therefore recorded with a printer. The endogenous respiratory rate from Pseudomonas sp. strain VB was determined in the reaction chamber without the addition of substrate. The oxygen consumption rate at the expenses of substrate was determined by subtracting endogenous respirations values from those obtained with substrate (exogenous). This was done to ascertain the possible intermediary metabolites in the biodegradation of 2–methoxyethanol by Pseudomonas sp. strain VB Determination of rate Limiting concentration of 2-methoxyethanol: The stoichiometry of 2-methoxyethanol and oxygen use by Pseudomonas sp strain VB was determined with Clarke-Type electrode. Briefly, 3ml of 50mM phosphate buffer (pH 7.2) was put in the reaction chamber at room temperature. The rate of oxygen consumption was measured with the intact resting cells (100 µ l) and limiting concentrations of 2–methoxyethanol. The oxygen reduction was recorded with compensation printer (Packard Model 613, Kpp and Zonen, Netherland). The endogenous rate oxygen uptake was measured without the addition of substrate. The exogenous rate of oxygen up take was corrected for the endogenous respiration rate. Measurement of 2-methoxyethanol Loss: In order to measure the biodegradation of bacterium isolate, Pseudomonas sp. strain VB previously described (Ekhaise, 2002), was inoculated in pure culture into mineral medium (Meyer and-Schlegel, 1983) containing 2-methoxyethanol as carbon and energy source. Samples withdrawn every 1 hr were analyzed for the microbial growth rate and the 2– methoxyethanol profiles as a qualitative assessment of 2-methoxyethanol utilization as a sole carbon and energy sources. An appropriate volume of culture fluid was withdrawn and centrifuged for 10min at 10,000xg. The resulting supernatant was subjected to the determination of 2-methoxyethanol concentration using gas chromatography, equipped with a flame ionization detector (FID), using a types Packed Column (Hayesap 80 – 10 mesh 2m long, inner diameter 2mm, model 430 from Packard, Netherlands), operated isothermally 180° C (oven) and 200° C (injection port and detection). Protein Determination: Biuret reagent was used to determine protein concentrations, following the method outlined by Daniels et al. (1994). Bovine serum albumin (BSA) served as the standard. Determination of bacterial cell dry weight: Dry weight of Pseudomonas sp strain VB was determine using a membrane filter was kept overnight at 80° C preweighed and cooled in the vacuum (ca/1h). The prewighed membrane filters were used to filer bacteria suspension (10ml) in a filtration chamber (Nalgeness, Rochester, USA). After filtration, the membranes were dried overnight at 80°C, cooled in the vacuum and then reweighed. The bacterial cell dry weight was determined as the difference in the membrane filer before and after filtration (Gerhardt et al., 1994). Chemicals: All the chemicals used were of analytical grade ( ≥ 99.9% pure). The water used was utrapure double-distilled water. 2-methoxyetanol, methanol, ethylene glycol, glycollate and glyoxylate were obtained from Fluks (Buchs, Switzerland), Merck (Darmstadt, Germany) and Sigma (Deisenhofen, Germany). RESULTS AND DISCUSSIONBiodegradation: In the biodegradation experiment, an increase in bacterial numbers shown in Fig. 1 and 2 was obtained as a result of growth and the corresponding utilization pf 2-methoxyethanol, by the Pseudomonas sp strain VB. Pseudomonas sp strain VB grew aerobically in mineral medium with 2-methoxyethanol (126.7mmol) with a doubling time of 1.739h. The growth rate of the bacterial isolate during 2-methoxyethanol utilization showed that protein content and cell dry weight were proportional during the logarithmic growth. When the cell dry weight is approximately 50% carbon (Madigan et al., 2000), there appears to be cellular assimilation of 2-methoxyethanol with an efficiency of about 50%. This finding aggresses with the report of Madigan et al. (2000). Aerobic transformation of 2 – methoxyethanol and hypothetical pathways intermediates: Studies of the metabolic pathway for 2-methoxyethanol by Pseudomonas sp. Strain VB yielded the data shown in Table 1. A significant oxygen uptake was observed with ethylene glycol, glycollate, glyoxylate and glucose while negligible oxygen consumption was observed with acetate and the intact resting cells could not consume oxygen at the expense of methanol (Table 2). This result suggests that, ethylene glycol, glycollate and glyoxylate are possible intermediate metabolites in the biodegrading of 2–methoxyethanol by Pseudomonas sp. strain VB. This finding supported the proposed degradative pathways in the following sequence: 2-methoxyethanol → ethylene glycol → glycollate → glyoxylate → oxalate → carbon dioxide under aerobic conditions. The result accounts for the utilization of the intermediate metabolites. The stoichiometry of 2-methoxyethanol biodegradation and oxygen uptake with isolate as shown in Table 2 above was calculated to be 1 to 1 ratio. The isolate utilizes 1 mol oxygen per mol of 2-methoxyethanol as compared to 4mol oxygen (O2) per 1mol 2 –methoxyethanol by total oxidation of 2. The biodegradation of 2-methoxyetanol under anaerobic conditions has been extensively investigated (Tanaka et al., 1986; Tanaka and Pfennig, 1988). A single bacterium isolate that can utilize 2 –methoxyethanol as growth source identified as Pseudomonas sp strain VB isolated from anaerobic sewage plant in Bayreuth/ Germany is here –methoxyethanol under aerobic conditions. The results therefore support the idea that intermediary metabolites are involved during biodegradation of 2-methoxyethanol under aerobic conditions in the Pseudomonas sp. strain VB considered. The outline biodegradative steps involved in the metabolism of 2-methoxyethanol by the new bacterium isolate (Fig.2) suggests that cultures of Pseudomonas sp. strain VB can mineralize 2-methoxyethanol as well as its proposed intermediary metabolites. C3H8O2 + 4O2 4H2O + 3CO2 (Total oxidation of 2-ME) Eqn 1 The transformation of 2-methoxyehanol seems to be constitutively expressed in Pseudomonas sp. stain VB. This view is not at variance with the report of Diekert (1992). The new bacterium strain failed to mineralize methanol, suggesting that it may be cleaved as methyl radical or methyl species as is the case under anaerobic conditions (Diekert, 1992). In support of this argument is the electron microscopy of the intracytoplasmic membrane examination of the cells, which revealed that the cell lacks the typical intraaacytoplasmic membrane arrangement of the type I and II methylotrophs (Ekhaise, 2002). Since methanol was proposed as an intermediary metabolite in the biodegradation of methoxyethanol under aerobic conditions, it can be assumed that the present isolate does not belong to the methylotriophs because of the absence of the special arrangement of the intracytroplasmic membrane of type I and II methylotrophs (Madigan et al., 2000). However, stoichiometrically the strain utilized one mole of oxygen per one mole of 2-methoxythanol as opposed to four mole of oxygen per one mole of 2-methoxyethanol for the total oxidation metabolism (Table 2). These findings therefore support the proposed degradative metabolic pathways of 2-methoxyethanol under aerobic conditions by Pseudomonas sp. Stain VB, which therefore accounts for its conversion to intermediary metabolites. Concern about on-going man-made pollution is stimulating efforts to investigate new productive bacterial strains and their biodegradative pathways for bioremediation purposes (Portrawtke et al., 1998). The new 2-methoxyethanol utilizing Pseudomonas sp strain VB represents a novel and promising tool for the rational design of optimized microorganisms able to mineralize highly recalcitrant pollutants like 2-methxyethaol. ACKNOWLEDGEMENTS This work was supported by grant from Deutsche Akademische Austauschsdient (DAAD) Bonn, Germany as part of my Ph.D research. I am grateful to Prof. Dr. O. Meyer for his useful advice and my wife and daughter for the moral support during the programme. REFERENCES
Copyright 2011- Journal of Applied Sciences and Environmental Management The following images related to this document are available:Photo images[ja11011t2.jpg] [ja11011f1.jpg] [ja11011t1.jpg] [ja11011f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}