 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
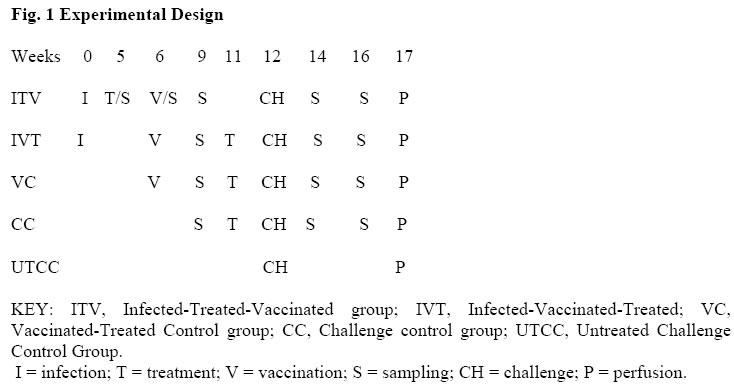
African Journal of Health Sciences, Vol. 13, No. 1-2, Jan-June, 2006, pp. 55-68 RESEARCH ARTICLE The effect of vaccinating S. mansoni–infected BALB/c mice either before or after treatment* Dorcas S. Yole¹*, Vincent O. Obanda², Kiio Kithome¹ and Horance Ochanda² 1.Institute of Primate Research, P.O. Box 24481, Karen Nairobi. 2. Zoology Department,
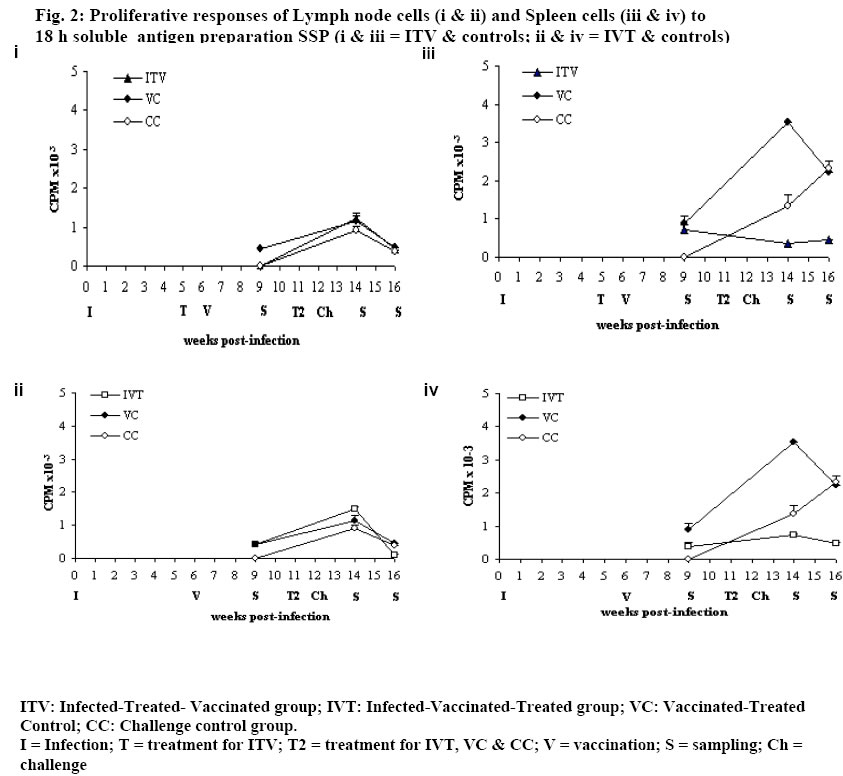
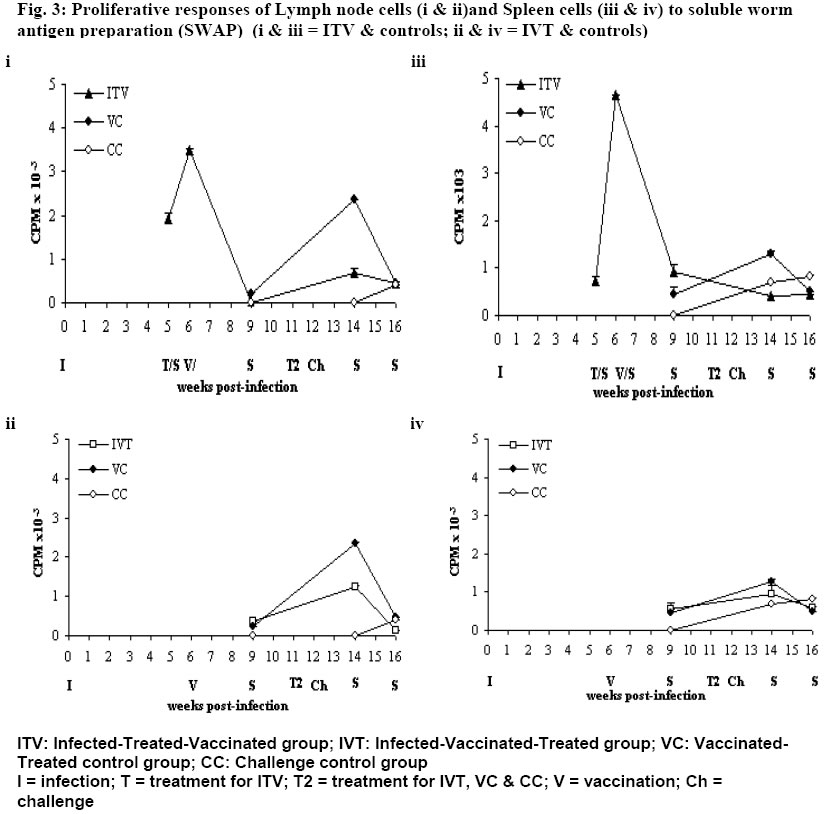
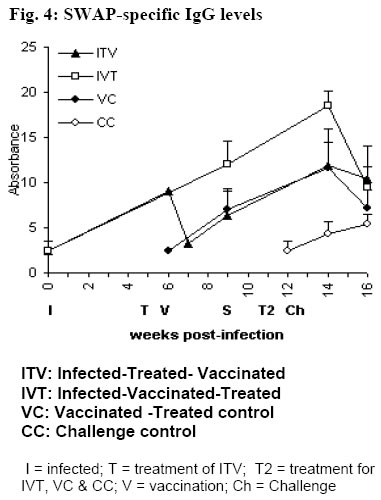
University of Nairobi, P.O. Box 30197, Nairobi, Kenya. Code Number: jh06009 SUMMARY In Schistosoma mansoni endemic areas, there are people with ongoing S. mansoni infection, others have been infected and treated while others have never been infected. What would happen if these different groups of people were vaccinated against S. mansoni? BALB/c mice were divided into five groups: Infected-Treated-Vaccinated; Infected-Vaccinated-Treated; Vaccinated-Treated Control; Challenge Control and Untreated challenge Control. Vaccination (500 20krad irradiated S. mansoni cercariae), Treatment (praziquantel), Infection and Challenge (150 S. mansoni cercariae) were carried out at specified times. Proliferation assay, Enzyme linked immunosorbent assay, gross pathology, histopathology and perfusion were performed. High protection levels were obtained in mice treated after vaccination: Vaccinated-Treated control, 96.5%; Infected-Vaccinated-Treated, 68.9%; and Infected-Treated-Vaccinated, 41%. A good correlation was obtained between proliferative responses and protective levels, implying cellular involvement in protection. Although all protected animals had high IgG levels, there was no strong correlation between the two. Specificity rather than amounts of IgG, seem more important in protection. Praziquantel seemed to boost protective immunity when administered after vaccination. Granuloma development and modulation in the two test groups was similar. It seems better to vaccinate infected patients before treatment, the ideal situation being vaccinating people who have not encountered S. mansoni . [Afr J Health Sci. 2006; 13:55-68] *Published online before print Introduction Schistosomiasis causes a chronic debilitating disease especially among the youth and children [1]. The global prevalence of the disease has remained approximately 200 million for decades [2], even in the presence of a safe and efficacious therapeutic drugs. Although chemotherapy remains the cornerstone of intervention, there is need for a more long-term approach, since the rapid re-infection demands frequent re-treatment that could eventually result to drug resistance [3,4]. Therefore it seems that the potential control of this disease will rely on multiple and integrated strategies, among which the modern tools of immunointerventions will play a major role. Based mainly on the murine challenge models, a number of candidate antigens have been proposed following immunizations with either irradiated cercariae or purified sub units [5,6]. These antigens usually demonstrate immunoreactivity with sera and peripheral blood mononuclear cells from infected patients [7]. Such studies have provided the justification for initiating vaccine development, with the aim of testing one or more candidate antigens in phase 1 clinical trials [8,5]. However, it is of paramount importance to evaluate any potential risk that may result during field clinical trials of a schistosomiasis vaccine. The reason being that in endemic areas, it is probable that there will be individuals who have undetected infections while others may have been infected and treated. The goal of the present study was to elucidate the effect of 20 krad radiation-attenuated vaccine when administered either before or after praziquantel treatment in S. mansoni infected BALB/c mice. Radiation-attenuated vaccine has given good protection levels in both murine and primate models [9-11]. Although the vaccine cannot be used in humans due to ethical reasons, it is useful in studying the immunological, parasitological and pathological responses of a working vaccine. Animals BALB/c mice, 8 weeks old (at the commencement of the experiment) were used in this study. They were bred at the Animal Resource Department of the Institute of Primate Research (IPR), Kenya. Mice were fed with nutrient pellets from Unga Feeds® Co., Kenya, and water supplied ad libitum. Materials and Methods Experimental design In this study, BALB/c mice were placed into five groups: Infected-Treated-Vaccinated (ITV), Infected-Vaccinated-Treated (IVT), Vaccinated- Treated Control (VC), Challenge Control (CC) and Untreated Challenge Control (UTCC). Mice in ITV were infected, treated with praziquantel, vaccinated and challenged. Mice in IVT were infected, vaccinated, treated with praziquantel and then challenged. VC mice were vaccinated, treated with praziquantel and then challenged. CC mice were treated with praziquantel 4 days before challenge infection. UTCC mice were infected with normal cercariae and not treated at all. Five mice were sampled per every sampling point. A schematic representation of the experimental design is given in Fig. 1 Parasite and Intermediate Host Schistosoma mansoni eggs were obtained from faecal material of infected olive baboons that were maintained at the Institute of Primate Research, IPR, Kenya. The eggs were hatched and 5-8 miracidia were used to infect each fresh water snails, Biomphalaria pfeifferi. The snails were maintained at the Snail Facility at IPR, Kenya. Preparation of the vaccine Normal cercariae were quantified and then irradiated by 20 krad emmited from caesium137 gamma source at the International Livestock Research Institute (ILRI), Kenya. Infection, Challenge and Vaccination Infected snails were shed and cercariae quantified for infection, challenge and for preparing radiation-attenuated vaccine at specified times. During infection, challenge and vaccination, mice were anaesthetized with a mixture of Rompun and Ketamine (Agrar, Holland) at 0.02ml/30g body weight via intraperitoneum. Mice for infection and challenge received 150 normal S. mansoni cercariae, while those for vaccination received 500 20 krad radiation-attenuated parasites each. Mouse abdomen was shaved and a metal ring 1cm in diameter was placed on the wet shaved area. Parasite suspension was placed in the metal ring. Parasites were given 30 minutes to penetrate. Treatment Praziquantel (Biltricide®, Bayer, Germany) tablets were ground into powder and then dissolved in distilled water. A dose of 450mg/kg/body weight was given orally to each mouse [12]. Two doses of the drug were given 48 h apart. Preparation of Antigens 18 hr soluble larval antigen (SSP) was prepared following artificial transformation of S. mansoni cercariae [13] and separation of heads from tails on a discontinuous Percoll gradient [14]. The schistosomules were cultured at 37oC, 5% CO2 in Complete medium [RMPI 1640, 0.2mg/ml Gentamycin, 1% glutamine (2MM); 1% 2mercaptoethanol (5x10-5 M), fortified with 10% foetal calf serum] under sterile conditions for 18 h. They were then pelleted and washed two times in PBS and sonicated (24 kHz, 16mm amplitude, 10 min) before centrifugation at 100,000 g for 1 h to obtain the soluble fraction. The protein content was determined [15] and the solution was UV-sterilized and stored at –70oC. Soluble worm antigen (SWAP) was prepared from 5-week-old worms recovered from baboons [10]. A commercially prepared Concanavalin A (Con A; Sigma Co.) was dissolved in sterile PBS to make a concentration of 1mg/ml. The solution was sterilized by Nalgene disposable filter (Nalgene Co., USA). Proliferation assay Spleen from each mouse was placed in sterile wire gauze, in a petri dish containing sterile Incomplete medium [RMPI 1640, 0.2mg/ml Gentamycin, 1% glutamine (2MM); 1% 2mercaptoethanol (5x10-5 M)]. A 10ml syringe piston was used to squash the organ. The dispersed cells were dispensed in 15ml tubes and topped up with Incomplete medium. The cell suspension was centrifuged twice at 450 g for 10 minutes, each time discarding the supernatant. The final pellet was re-suspended in 4 ml of Complete medium. Lymphocyte viability was determined by Trypan blue exclusion test. Lymphocytes were counted in a haemocytometer chamber. Cells for the culture were made up to 3x106 cells/ml in Complete medium. Lymph node cells were prepared by teasing the organ using sterile forceps in petri dish containing sterile Incomplete medium. The cells were prepared for culture based on the procedure used for spleen cells. Spleen cells and lymph node cells were cultured in flat-bottomed 96-well microtitre plates (Nunclon, Denmark). Duplicate wells were set for each regime, each well containing 3x105 viable cells. Negative control had the medium and cells only while positive control had 1 µg/well of Con A. The test wells of each plate contained 1 µg/well of soluble worm antigen preparation (SWAP) or 0.5 µg/well of 18 hr soluble larval antigen preparation (SSP). The total volume of culture medium per well was made up to 200 µl. The plates were incubated at 37oC, in the presence of 5% carbon dioxide. After either 48 h or 72 h for Con A and for other set-ups, respectively, the cells were pulsed with 20 µl of 18.5 MBq tritiated thymidine (3H-thymidine; Amersham Pharmacia, UK). After 18 h, the cells were harvested on microfibre filter using Filtermate Harvester (Packard Bioscience Co. USA). The incorporated label was measured by liquid scintillation counting. Results were expressed as differential counts, ∆ CPM, that is CPM of Con A or antigen-stimulated cultures – CPM of negative controls. (CPM means counts per minute). Antibody assay The SWAP-specific ELISA was performed by coating Immunolon 4 plates (Dynatech, USA) with 10 µg/ml of SWAP in carbonate bicarbonate buffer (pH of 9.6) overnight at 4oC. Plates were subsequently blocked with PBS plus 0.05% Tween 20 (Fisher Chemicals, USA) and 3% BSA for I h at 37oC. After washing, a 1:4 dilution of mice sera was added and incubated at 37oC for 2 h. Plates were washed again and goat anti-mouse IgG conjugated to horse-radish peroxidase was added and incubated for 1 h at 37oC. Plates were washed again, substrate (TMB micro well peroxidase substrate, Kirkegaard & Perry Labs, USA) was added, and optical density (OD) determined at 630 nm. Perfusion and worm recovery Based on the modified method of Smithers and Terry [16], mice were anaesthetised and hepatic portal vein incised. Perfusion needle containing perfusion fluid (0.85% Sodium chloride and 1.5% Sodium citrate) was inserted on the left ventricle of the heart and perfusion carried out until the liver, lower limbs and mesenteries were clear. The perfusate was collected in plastic container and transferred in a urine jar to settle. The supernatant was sucked out and worms recovered [10]. The mean and percentage worm reduction of adult worms recovered for each group was calculated. Worm maturation was also calculated for the control groups. Gross pathology and histopathology Gross pathology of the liver and the mesenteric lymph nodes was observed. The observations considered in the liver included the colour, size and presence of granulomas. Representative biopsy samples of the liver were fixed immediately in 10% buffered formalin for at least two weeks. The tissues were cleared in toluene, infiltrated in hot paraffin and embedded on tissue-embedding paraffin wax (Sherwood Medical Co. USA). The tissues were sectioned serially at 6 microtomes using a Rotary microtome (Leitz, Germany). The thin tissue sections were mounted on glass slides and stained with haematoxylin and eosin. The tissue sections were observed under light microscope and only granulomas containing an ovum in the centre were enumerated and measured at 100x magnification using calibrated ocular micrometer. The vertical and horizontal diameters of granulomas with a visible centrally placed schistosome egg were measured using the ocular micrometer [12]. The averages of the horizontal and vertical diameter were taken to be the diameter of the granulomas. Ten granulomas were measured for each mouse. Statistical analysis Analysis of variance (ANOVA), Kruskal Wallis test and the Student's t-test, were used in data analysis. P values <0.05 were considered significant. Results Lymphocyte Proliferative responses Lymph node (LN) cells responses to SSP are shown in Fig. 2 (i) and (ii). The three vaccinated groups: Infected-Treated-Vaccinated (ITV), Infected-Vaccinated-Treated (IVT) and Vaccinated-Treated Control (VC) groups had similar responses to SSP. They had either low or background responses 3 weeks post-vaccination (wk pv). The responses were boosted by challenge infection as demonstrated by a peak 2 weeks post-challenge (wk pc). However, at 4 wk pc, the responses were either low or at background levels. The Challenge Control (CC) had substantial response 2 wk pc, but the level had gone down by 4 wk pc. Spleen Cells responses to SSP are shown in Fig. 2 (iii) and (iv). Although the trend for spleen cells responses for mice IVT and the VC was similar, the responses for the former group were much lower (Fig 2.iv). The two groups had noticeable responses 3 wk pv, a peak at 2 wk pc and a decline 4 wk pc. Spleen cells of ITV had remarkable response at 3 wk pv, which declined slightly 2 wk pc and remained almost at the same level at week 4 wk pc (Fig. 2.iii). CC had high response at 2 wk pc. The responses were still going up by 4 wk pc. Proliferative responses to SWAP: Lymph node cells responses to SWAP are shown in Fig. 3 (i) and (ii). LN cells of IVT and the VC demonstrated similar responses to SWAP, although VC had higher response (Fig. 3.iii). Both had low responses at 3 wk pv. They attained a peak at 2 wk pc with the peak being higher for VC. Unlike the VC, IVT had a background response by 4 wk pc. ITV had high responses at 5 wk post-infection (pi; Fig. 3.i). The first peak was observed 1 wk after treatment, at the time when large amounts of antigen were being released from the dying worms. However, at 3 wk pv, no response was observed. Two weeks after challenge, there was a remarkable response, which was only slightly decreased 4 wk pc. The Challenge Control group had a response recorded only at wk 4 wk pc at the time when worms start maturing. Spleen cells responses to SWAP are shown in Fig. 3 (iii) and (iv). Spleen cells of IVT and VC had similar responses, with a remarkable response at 3 wk pv, a peak at 2 wk pc and a decline by 4 wk pc Spleen cells of ITV had reasonable responses at 5 wk pi and a peak was obtained 1 wk after treatment. The response had decreased by 3 wk pv, and was lower 2 wk pc, though it remained remarkable. The response remained more-or-less at the same level by 4 wk pc. The challenge control animals had a high response by 2 wk pc, and by 4 wk pc, a slight elevation was noticeable. IgG antibody responses SWAP-specific IgG responses of the four groups of mice are shown in Fig. 4. IVT had a steady increase of IgG from infection up to a peak at 2 wk pc, followed by a drop at 4 wk pc. VC had a Warm Maturation Worm maturation from Challenge Control (CC) and Untreated Challenge Control (UTCC) was calculated as shown below;
CC, which had received treatment prior infection, had a mean burden of 17 worms and worm maturation of 11.3%. UTCC, which was just infected without being treated, had a mean Burden of 29 worms and worm maturation of 19.3% Worm resistance When worm burdens for the vaccinated groups were subjected to ANOVA, there was a significant difference (F 4,16 = 11.307, P = 0.000) between the groups vaccinated before treatment (IVT and VC) and the group treated before vaccination (ITV). The above results were confirmed by the Kruskal Wallis test (X2 = 16.664, P = 0.002), Post hoc (Turkey) analysis. The mean difference was significant at the 0.05 level. There was no significant difference between IVT and VC. Worm resistance for ITV, IVT and VC was calculated using the formula shown below.
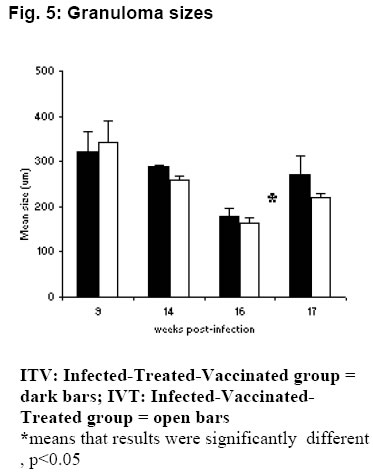
The highest worm resistance was attained by VC, 96.5% followed by IVT with 68.9% and finally ITV with 41%. Gross pathology At 9 wk pi (3 wk pv), the surfaces of the liver lobes from ITV and IVT were covered entirely by whitish /creamy spots (granulomas). The usually smooth liver surface was replaced with rugged surface. Moreover, the liver appeared pale as opposed to the red-pink hue in normal livers. The organ was enlarged and edematous. At 14 wk pi (2 wk pc) the liver in these two groups showed no gross pathological differences from the ones observed at 9 wk pi (3 wk pv), except that the livers had become dark in appearance. At 16 wk pi (4 wk pc), livers from mice in group ITV and IVT were darker than previously noticed. Granulomatous spots were numerous and covered every part of the liver lobes. The Granulomas appeared to be protruding slightly above the liver capsule. The livers were gritty to touch and creamy patches were also observed at the edges of the liver lobes in some mice. The liver was prominently enlarged. At 17 wk pi (5 wk pc), gross pathology of the liver of these two groups showed no marked difference from the observations made at week 16 wk pi (4 wk pc). Throughout the sampling period, livers from the VC and CC, had the texture, size and colour of normal liver. There were no observable changes. The livers lacked visible granulomas and retained the smooth texture, sharp edges and pink-red hue. There was no pronounced liver enlargement or oedema. Enlarged mesenteric lymph nodes were also observed in different mice groups at different time points. Enlargement of the mesenteric lymph nodes of ITV was observed at 9 wk pi (3 wk pv) and the size remained more or less the same from 2 wk pc to week 5 wk pc. IVT mesenteric LN were enlarged at 9 wk pi (3 wk pv) and became more enlarged and inflamed as from 14 wk pi (2 wk pc) up to 17 wk pi (5 wk pc). VC mesenteric LN were enlarged from 14 wk pi (2 wk pc). CC mesenteric lymph node showed slight enlargement as from 16 wk pi (4 wk pc). Histopathological findings Granuloma diameters are shown in Fig. 5. Peak liver granuloma size was observed 9 wk pi (3 wk pv) in both ITV and IVT. The granulomas were large and florid each with a conspicuous egg at the centre. Hepatic granulomas from mice in ITV, did not show significant granuloma size reduction between 9 wk pi (3 wk pv) and 14 wk pi (2 wk pc), whereas, IVT showed some reduction from 9 wk pi (3 wk pv) through to 14 wk pi (4 wk pc). At week 16 pi (4 wk pc), both ITV and IVT had significantly smaller granulomas, (P <0.05), as compared to those of 9 wk pi (3 pv). At 17 wk pi, granuloma sizes of ITV and IVT had significantly increased as compared to 16 wk pi (P <0.05). The VC and CC lacked granulomas with centrally placed egg. However, CC had few patches of inflammatory foci. Periportal fibrosis and periportal cellular infiltration were observed in some liver tissues from mice in both ITV and IVT. Periportal fibrosis surrounded the portal vein, while diffuse cellular infiltration were observed around the bile ducts. In such tissues, granulomatous foci were scanty or absent completely. Adult worms embolized on the portal veins were observed in the liver sections from mice in these two groups only. The worms were observed by week 9 pi (3 wk pv) in ITV, while in IVT they were observed by 14 wk pi (2 wk pc). Such portal veins were heavily surrounded with thick layer of fibrotic material on the inner wall, while on the outside, heavy cellular infiltration were very conspicuous. Coalescing granulomas surrounding two or three schistosome eggs were also observed in livers from the two groups. Intestinal granulomas were only observed in mice from ITV and IVT. These granulomas were few but diffuse in size. They were found mostly within the serosal surface of the intestine. Discussion Proliferative responses Antigen specific lymphocyte proliferation, the assay commonly used for assessing cellular responses after immunisation, was used in this study. The presence of lymphocytes that have memory to a specific antigen are detected by their subsequent in vitro proliferation when reexposed to the same antigen. Although, this assay does not reveal the functional capabilities of the responding lymphocytes, it does indicate that cell are able to recognise the particular antigen that had activated them previously. Given the supposed involvement of lymphocytes in immune elimination of challenge schistosomula from vaccinated mice [17, 18] it might be anticipated that successful vaccination would preferably stimulate the proliferation of T cells in particular draining lymph nodes. Both lymph node and spleen cells from the Vaccination-Treated Control group demonstrated the highest proliferative responses to schistosome antigens as compared to the other three groups. Notable responses were observed 3 weeks after vaccination; a peak response was recorded 2 weeks after challenge, then a decline at 4 weeks post-challenge. This finding is in agreement with other studies in which proliferative responses of lymphocytes to schistosomal antigens were found to peak during the first two weeks after vaccination, waning after the fourth [19, 20]. The attenuated parasites release significant amounts of antigenic material during their passage through the skin, lymph nodes and lungs where they eventually die [19, 20]. Responses to SWAP at week 3 postvaccination were substantial, a time when the vaccinating parasites were still at the larval stages. This response to SWAP could be due to the extensive cross reactivity between worm antigens and schistosomula antigens [21]. Proliferative responses to schistosome antigens of lymph node and spleen cells from Infected-Vaccinated-Treated mice had similar patterns to the Vaccination-Treated Control groups. However, at almost all points, the responses were lower than those of the Vaccination-Treated Control group. Treating this group just before challenge resulted in large amounts of antigens being released from the dead worms (infection parasites). Additional amount of antigen was released by the challenge parasites themselves. The large amount of antigen could have resulted in anergic responses, hence the lower responses recorded in this group as compared to the Vaccination Control group. Worms killed by praziquantel introduce a large amount of antigens directly into the blood stream and their persistence may cause overstimulation of lymphocyte clones, leading to suppression of proliferative response. Moreover, the presence of high concentration of antigens in circulation may favour direct interaction of antigens with lymphocytes without the participation of antigen presenting cells (APCs) and thus cause unresponsiveness [22]. On the contrary, at the time when the Vaccinated-Treated Control mice were treated at week 5 post-vaccination, all the vaccinating parasites had died [23] hence no extra amounts of antigen were released from the parasites in this group which would have resulted to an anergic effect. In the Infected-Treated-Vaccinated mice, responses of both lymph node and spleen cells to SWAP, before vaccination were very interesting. High proliferative responses were observed at week 5 post-infection. By week 5, worms have matured and started laying eggs; antigens from both adult worms and eggs were responsible for the high proliferative responses. In this group, mice were also treated at week 5, and by week 6 peak responses were attained which were higher than any other responses observed in the entire sampling period. This was due the immediate response of antigens released from the dead worms. However by week 3 post-vaccination, the responses had gone down. This could be attributed to anergic effect of the large amounts of antigens from dead worms. Antigens released from vaccinating parasites most probably encountered unresponsive lymphocytes resulting into the not so dramatic response observed 3 weeks after vaccination. This lymphocyte unresponsiveness seems to have remained even after challenge. In the Challenge Control group, responses to the schistosome antigens were recorded at week 2 and 4 post-infection for both cell types, except for lymph node cells responses to SWAP which were only observed at week 4 postinfection. This is in agreement with previous observation [20]. IgG responses The IgG isotype, particularly IgG1, seems to be protective and may be synergistically enhanced by IgM [24]. The Infected-Vaccinated-Treated group had the highest IgG levels throughout the experiment except at week 4 post-challenge. Vaccination, treatment and challenge seem to have boosted the effect of infection, resulting to a steady increase of SWAP-specific IgG to a peak at week 2 post-challenge. The drop at week 4 post-challenge could be attributed to lower antigen load as a result of few challenge parasites, compounded by death of challenge parasites as a result of vaccination [23]. The Vaccinated-Treated Control group had a similar pattern to the Infected-Vaccinated-Treated group from week 3 post-vaccination, except that the levels were much lower. As expected, vaccination resulted in elevated IgG levels that were boosted by challenge. IgG titres are enhanced by repeated vaccinations with irradiated cercariae, or after challenge infection with normal cercariae [25]. The levels were much lower as compared to the Infected-Vaccinated-Treated group most probably because the Vaccinated-Treated Control group encountered the parasite only twice while the other group had more encounters with parasite antigen. Among the three vaccinated groups, the Vaccinated-Treated Control group had the lowest level of IgG at week 4 post-challenge, most probably due lower parasite numbers at this time as a result of parasite killing by the vaccine. The Infected-Treated-Vaccinated group had high IgG levels by week 6 post-infection, most probably as a result of large amount of antigens released from the dying worms. However, by week 7, the levels had gone down because of clearance of antigens from dead worms from circulation. Vaccination and challenge increased the antigenic load in the circulation, and at week 3 post-vaccination and week 2 post-challenge, both this group and the challenge control group had similar IgG levels. On the other hand, at week 4 post-challenge, this group recorded the highest level of IgG among the three vaccinated groups. The Challenge Control group SWAPspecific IgG levels were evident at week 2 postinfection and had gone up by week 4 postinfection, but not as high as the other three groups. This could be attributed to the low antigenic load as a result of one infection only. A similar trend was reported for baboons infected with. S. mansoni [10]. Worm maturation and resistance The Challenge Control group that had been given oral dose of praziquantel 4 days prior to challenge had a lower worm maturation (11.3%) as compared to the Challenge Control group, which did not receive any treatment prior to challenge (19.3%). The results show that the drug might have sensitized the immune system of the treated Challenge Control against the penetrating parasites. Praziquantel metabolites may have triggered non-specific antibody responses and/or the presence of the schistosomula may have provoked antigen dependent cellular cytotoxicity (ADCC). The surface of the newly transformed schistosomula of S. mansoni is highly antigenic and the larvae are very susceptible to ADCC [26]. Plasma elimination half-life for the unchanged drug is 1-1.5 h and 4-5 h for metabolites after a single dose, ruling out the effect of drug residue on the challenge parasites. There was a significant difference between groups vaccinated before treatment (Infected-Vaccinated-Treated group, and the Vaccinated-Treated control group) and the group vaccinated after treatment (Infected-Treated-Vaccinated group), using both the ANOVA and Kruskal Wallis test. However, no significant difference was found between the Infected-Vaccinated-Treated group, and the Vaccinated-Treated control group. This points to the fact that vaccination before or after treatment affects the worm burden. The Vaccinated-Treated control group had the highest protection level of protection, 96.5%. A protection level of 60-70% has been reported for C57BL/6 mice [10]. The high protection level could be as a result of a different strain of mice, which is more resistant, and or to treatment with praziquantel after vaccination, which could have enhanced the immune protective mechanism. Infected-Vaccinated-Treated mice had the second highest level of protection, 68.9% which is in the normal range of reported protection for mice [10]. The Infected-Treated-Vaccinated group had the lowest protection level of 41%. It is obvious that mice which were vaccinated before praziquantel treatment (Infected-Vaccinated-Treated and Vaccinated-Treated Control groups) had higher resistance against challenge parasites as compared to mice that were treated with praziquantel before vaccination (Infected-Treated-Vaccinated group). The high anti-worm activity in Infected-Vaccinated-Treated and Vaccinated-Treated Control groups could be attributed to the synergistic effects of praziquantel and the vaccine. The reason being that the effectiveness of praziquantel against S. mansoni infections is increased in vaccinated mice [27]. This explains why synergistic effect was not substantial in Infected-Treated-Vaccinated group, which was treated before vaccination. This observation is consistent with other experimental findings that showed the enhanced efficacy of the drug in immune sensitised hosts [28, 29]. The protection levels in these three groups seem to correlate very well with the proliferative responses. The Vaccinated-Treated Control group had the best proliferative responses after vaccination, followed by Infected-Vaccinated-Treated group and then the Infected-Treated-Vaccinated group. This points to the involvement of cellular immunity in protection. Although the presence of antibodies seems essential for protection, there is no consistent association between antibody levels and level of protection. It seems that antibody specificity rather than quantity is more relevant to protective immunity. However, the level of antibody seems to correlate very well with the amount of antigen in the circulation. This was demonstrated by: the peak at week 6 and sharp drop at week 7 of the Infected-Treated-Vaccinated group, corresponding to high antigenic load from dead worms, and its clearance, respectively; consistently high levels of IgG in Infected-Vaccinated-Treated group, which had high antigenic loads throughout the experiment; and the correlation of level of IgG at week 4 post-challenge for the three vaccinated groups and number of worms recovered after perfusion. Pathology Mesenteric lymph nodes that drain the duodenum, caecum, colon and ileum were enlarged in all the mice groups. It has been observed that large amounts of schistosome antigens are conveyed to the mesenteric lymph nodes and become sequestered in the medullary cords [30]. Peak hepatic granulomas were measured from mice in both the Infected-Treated-Vaccinated and Infected-Vaccinated-Treated groups at week 9 weeks post-infection. This finding was in agreement with the established period of peak (acute) granulomas in mice. Granuloma peaks in size by 8-10 weeks after infection [31]. There was significant reduction (P<0.05) in hepatic granuloma size in Infected-Vaccinated-Treated and Infected-Treated-Vaccinated groups by week 16 post-infection. This was in accordance to other studies where hepatic granulomas begin to reduce in size and cellularity after about 10 weeks post-infection such that by 16 - 20 weeks post-infection, the granulomas are about half the size of acute granuloma [32]. This process is termed as immunomodulation. However, there was an unusual observation in mice from the Infected-Treated-Vaccinated and Infected-Vaccinated-Treated groups at week 17 post-infection. The hepatic granulomas from these mice showed significant (P<0.05) increase in size. At week 17 post-infection, parasite antigens and soluble egg antigens were being released continuously into host circulation from the challenge parasites. Consequently, the subsequent cellular infiltration caused by increased cross-reactivity of antigens from challenge schistosomula and schistosome eggs, may have caused 'old granulomas' to appear larger. Interchanging treatment and vaccination did not seem to unduly affect granuloma progression. The formation of granulomas around schistosome eggs in the liver and the intestine is the major cause of pathology in schistosome infections. The live miracidia within each egg secretes antigenic materials through ultrasmicroscopic pores in the shell [33]. These antigens, continually released for 2 to 4 weeks, induce host sensitisation and recruitment of macrophages, lymphocytes, giant cells, fibroblasts and numerous eosinophils to comprise the host granulomatous response [33]. Granulomas are thought to be host protective, as they wall-off toxic egg products, such as hepatotoxic antigen omega-1 [34], which would otherwise kill the host, (and consequently, the parasite too). In mice infected with S. mansoni, egg deposition begins about 4 to 5 weeks after infection with the first detectable granulomas present by about 6 weeks. The granulomas were large due to peak cellular responses in the host because of schistosomula surface antigens and worm tegumental antigens cross-reacting to egg antigens [21]. The presence of worms in the liver could be attributed to praziquantel treatment. The effects of the drug could have caused worms to be dislodged and then be swept via the portal system to the liver. Conclusion In conclusion, administering vaccine before praziquantel treatment gave better protective results as compared to administering vaccine after treatment. The best results were obtained when the vaccine was administered to naïve mice. There was a correlation between the protective levels and proliferative responses implicating cellular involvement in protection. Although the level of IgG seemed to correlate with antigenic load, there was no consistent association between antibody levels and the level of protection. Praziquantel seemed to influence protective immunity positively when administered after vaccination, but not before. It also seemed to have protective effect on mice treated before infection. Granuloma progression in Infected-Treated-Vaccinated and Infected-Vaccinated-Treated groups was rather similar. It followed the normal process of increase in size followed by modulation, except that the challenge infection resulted into increase in size in modulating granulomas in both groups. This work shows that it is crucial to know the state of vaccination target group before and during administration of a vaccine, as varying results may be obtained depending on the status of the individual before vaccination. Acknowledgement We would like to thank Sammy Kisara, Simon Kiarie, Fred Nyundo and Meshack Rakamba for technical assistance. References
Copyright 2006 - African Forum for Health Sciences The following images related to this document are available:Photo images[jh06009f2.jpg] [jh06009f5.jpg] [jh06009f1.jpg] [jh06009f3.jpg] [jh06009f4.jpg] |
| |||||||||

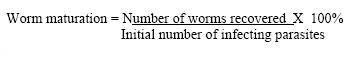
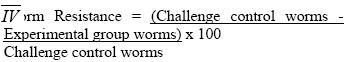
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}