 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
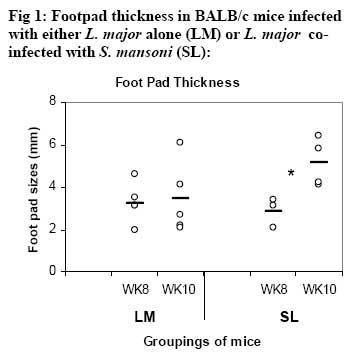
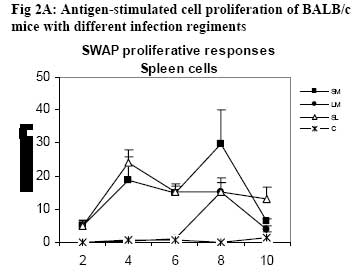
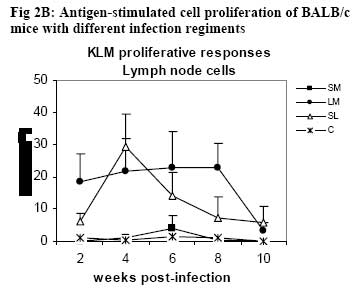
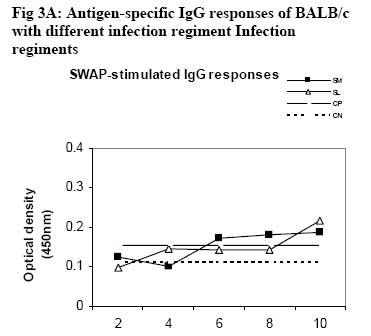
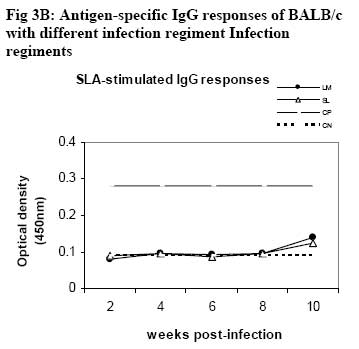
African Journal of Health Sciences, Vol. 14, No. 1-2, Jan-June, 2007, pp. 80-85 Studies on the interaction of Schistosoma mansoni and Leishmania major in experimentally infected Balb/c miceDorcas S. Yole¹*, Kennedy T. Shamala², Kiio Kithome¹, Michael M. Gicheru¹ 1. Institute of Primate Research, National Museums of Kenya, P.O. Box 24481, Karen, Nairobi, Kenya. Code Number: jh07011 SUMMARYSchistosoma mansoni and Leishmania major are important tropical human parasites. It is crucial to know the effect of the two infecting man concurrently. Two groups of BALB/c mice were infected with each of the parasites separately; another group was co-infected with both parasites and there was a naïve control. Draining lymph node and spleen cells from mice infected with either of the parasites showed high proliferative responses to their specific parasite antigen. However, crossreactivity occurred between S. mansoni and L. major. Spleen and Lymph node cells from co-infected group demonstrated high and sustained proliferative responses to schistosome soluble worm antigen preparation and killed Leishmania major antigen, respectively. There was high and sustained IgG levels for both the single and coinfected groups. At 10 weeks post-infection, co-infected mice had significantly larger nodules than mice with L. major infection alone. However, co-infected animals had less severe liver pathology and less enlarged mesenteric lymph nodes than those infected with S. mansoni only. This work shows that co-infection results in two different outcomes: protection against S. mansoni and exacerbated pathogy in L. major. We suggest that cellular responses possibly protect against S. mansoni, while high IgG levels lead to exacerbated L. major response. Introduction Schistosomiasis and leishmaniasis are two parasitic diseases associated with great human suffering in the endemic areas. The two diseases are widespread and double infection is not an uncommon feature [1, 2]. Interestingly, some infections with Schistosoma (a parasitic helminth) and Leishmania (a parasitic protozoa) share many clinical manifestations [1] thus confusing diagnosis of either of infections. Schistosoma mansoni infection localizes in the liver and the guts inducing formation of granuloma associated with Th2 dominant immune response while Leishmania major infection results into localized cutaneous lesions associated with Th1 response in susceptible mouse models [3,4]. However in BALB/c mouse model, L. major infection is associated with a non-healing phenotype, characterized with Th2 response [3,5,6,7]. A concomitant infection (co-infection) of S. mansoni and L. major in BALB/c is interesting in that Th2 immune response is protective for S. mansoni infection while the same responses is associated with disease exacerbation in L. major infection. It is not yet clear how concomitant infection of S. mansoni and L. major in BALB/c would influence pathogenesis of either of the infections. Such information is useful and may shed light on pathogenesis of S. mansoni or L. major in polarized Th2 environment. We report here results of a study in which we compared the pathogenesis and immune responses of a concomitant infection of S. mansoni and L. major against respective single parasite infection in the BALB/c mouse model. Material and Methods Hosts and parasites BALB/c mice bred at the Institute of Primate Research (IPR) were used in the experiments. A Kenyan isolate of S. mansoni, originally derived from infected human and maintained under laboratory conditions in Biomphalaria pfeifferi and baboons, was used to infect mice. Leishmania major strain NLB-144 originally isolated from the sandfly, Phlebotomous duboscqi, in Baringo District , Kenya, and maintained in BALB/c mice by serial subcutaneous passages was used in the experiment. Preparation of parasite antigens Schistosome soluble worm antigen (SWAP) was prepared from 6-week-old S. mansoni worms recovered from baboons. The worms were washed twice in PBS, and sonicated (24 kHz, 16mm amplitude, 10 min). The suspension was then centrifuged at 1x105 g for 1 h, at 4°C to obtain the soluble protein fraction [8]. The protein content was determined using the Bradford method [9]. The antigen was sterilized by exposure to UV light for 10 min, aliquoted and stored at –20oC. Leishmania major antigens (KLM) for proliferation assays was prepared from formalin fixed L. major promastigotes as described by Olobo [10]. Briefly the parasites were obtained from cultures at the stationary phase. The parasites were washed, enumerated and fixed in 1% formal saline for 1 hr and washed again. The parasites were then resuspended in sterile PBS at a concentration of 5x 108 promastigotes/ml and stored in –70oC until use. Soluble Leishmania antigen (SLA) for antibody ELISA was prepared as described by Gicheru [11]. Infections of BALB/c mice with parasitesExperimental mice were divided into four groups of 25 mice each. Each mouse of Group 1 received 150 S. mansoni cercariae percutaneously via abdominal skin using the ring method [12]. Group 2 mice were each inoculated with a total of 3.0 x 106 promastigotes of L. major in 50 �l PBS in the hind footpad. Group 3 mice were each infected with both S. mansoni and L. major. Group 4 mice were not infected with any parasite and served as naive controls. Sampling proceduresEvery two weeks following infection of mice with parasites, sera and lymphocytes were harvested from five mice in each group. The five mice from each group were euthanized, the thoracic cavity opened, blood obtained by cardiac puncture and serum prepared for the antibody enzyme linked immunorsorbent assay (ELISA). Inguinal and poplitual lymph nodes (draining sites for S. mansoni and L. major, respectively) and spleens were obtained for cell preparation. Measurements of lesion sizes on footpad were taken in L. major infected and coinfected mice groups at week 8 and 10. Similarly, pathological changes in liver and mesenteric lymph nodes in S. mansoni infected groups were observed at week 8 and 10. Proliferative responses of mouse spleen cells to SWAPSpleens were sampled individually from each mouse, for all the four groups of mice. Spleen was removed from each euthanized mouse and transferred to a petri dish containing sterile incomplete RPMI 1640 medium [RPMI 1640 (GIBCO, Paisley, United Kingdom) containing 100�g gentamycin (GIBCO), 2mM L-glutamine (Sigma, Laboratories, United kingdom) and 0.05mM 2-mercapto-ethanol (Sigma)]. Using a 10ml syringe piston, the spleen was squashed through a fine wire mesh. Each cell suspension was dispersed with a sterile Pasteur pipette, sucked and dispensed into a 15ml sterile test tube. The cells were washed twice by centrifugation at 450 g for 10 minutes at room temperature and enumerated. Their concentration was made up to 3x106 cells in 1 ml of complete RPMI 1640 medium (incomplete medium fortified with 10% foetal calf serum, Flow laboratories, U.K.) Triplicate cultures were set up in 96-well microtitres plates at a density of 3 x 105 cells/ 200ul medium/well of lymph node and spleen cells. Negative wells contained medium alone, positive control wells 1 µg Concanavalin A (Con A), and test wells 10 µg of SWAP. After culturing the cells for 48 h (Con A) or 72 h (antigens) at 37 oC, in a humidified incubator supplied with 5% CO2, 18.5 kBq [3H]thymidine (specific activity 185 GBq; Amersham) was added to each well. After 18 h, cells were harvested and the incorporated label measured by liquid scintillation counting. Results were expressed as differential counts, ∆, counts per minute (cpm; that is, cpm of Con A or antigen-stimulated cultures minus cpm of negative control). Proliferation responses of lymph node cells to L. major antigen Lymph nodes were obtained from each mouse in all the four groups of mice. Lymph nodes from each mouse were transferred to sterile petri dishes. Cells were obtained by teasing the node with forceps in incomplete RPMI. The cells were washed and made up to 3x106 cells/ml in complete medium. Triplicate culture wells were set up as described for spleen but the test antigen was 5 x 105 promastigotes per well. Setting negative and positive wells was done as described for spleen cells, so were culturing, harvesting and scintillation counting. Measurement of specific IgG levelsELISA plates (Dynatech, UK) were coated overnight with 50 µl of either SWAP (20ug/ml) or SLA (10ug/ml). Non-specific binding was blocked by incubation with 100 µl of 3% BSA for I h at 37oC. Mice sera from each group was pooled, and then serially diluted at a factor of 1:5. Negative control was set up using sera of naïve animals and positive controls using sera from mice infected with either S. mansoni or L. major. Each well contained 100 µl of diluted serum. IgG binding was detected using 100 µl of rabbit anti-mouse horseradish peroxidase conjugate (1/5000; Sigma, UK). All the incubations were for 1 h with appropriate washing steps. The substrate, 0-phenylenediamine was added to each well at 100 µl/well. The colour development was read at 450 nm after 30 min and 1 h. Statistical analysisStudent’s t-test was used to compare means between the groups. The confidence level was taken as 95%. Results Pathology associated with concomitant L. major and S. mansoni infections Footpad thickness of mice with concomitant L. major and S. mansoni infection were similar to those of mice with L. major infection alone, up to week 8 post-infection (Fig. 1). However by week 10 postinfection the footpad thickness of mice co-infected with both L. major and S. mansoni were significantly larger than those of mice infected with L. major alone (Fig. 1; P <0.05). For Fig 1, open circle represent thickness for an individual mouse in the specified group; * implies significant difference (p<0.05) Liver pathology was examined in mice infected with S. mansoni only, and in S. mansoni mice co-infected with L. major at week 8 and 10. The liver texture and granuloma numbers were assessed using a subjective score of “mild”, “moderate” or “severe” depending on severity on granulation. Overall, liver pathology in S. mansoni infected mice was considered to be moderate to severe, and 60% of the mice in this group had died by week 10. The dead mice had severe pathology. On the other hand, mice with concomitant L. major and S. mansoni infections had mild to moderate pathology based on the assessment criteria and no mortality was observed in these mice. As expected, severity of pathology increased from week 8 to week 10 in both groups. With respect to mesenteric lymph nodes, these were categorized as small, medium or large depending on their degree of swelling. Mice with S. mansoni infection only had medium mesenteric lymph nodes at week 8 but by week 10, they were large. The situation was however different in the mice co-infected with S. mansoni and L major. At week 8, all the mice in this group had small mesenteric lymph nodes, and by week 10, all were medium size, suggesting that the infection in mice with concomitant S. mansoni and L. major was less severe. Proliferative responsesIn general, cellular proliferative responses to Con A were greater than those observed for either S. mansoni or L. major antigens (data not shown). This is expected since Con A is a non-specific lymphocyte proliferation stimulator. When S. mansoni soluble worm antigen (SWAP) was used to stimulate spleen cells, proliferative responses greater than the background (negative control) responses were observed at all time points for cells from S. mansoni infected and for S. mansoni-L. major co-infected mice, and no significant differences were observed in responses between the two groups (P>0.05; Fig. 2A ). Low responses were observed at week 2, with a definite increase in response at week 4, but the responses remained the same up to week 8. Surprisingly, a response greater than the background was observed for cells from L. major infected mice, at week 8. Proliferative response to L. major antigen (KLM) was tested in lymphocytes obtained from draining lymph nodes. Responses from L. major infected or co-infected groups were greater than the background throughout the period of experimentation. However, mice infected with L. major alone gave a stronger and sustained response than the mice co-infected with S. mansoni and L. major (Fig. 2B). Responses to KLM antigen in the mice infected with S. mansoni only were greater than the background responses at week 6. Both spleen and lymph node cells from the control group showed background responses. IgG responsesFollowing infection, mice were tested for IgG responses to S. mansoni (SWAP) or L. major (SLA) antigens every two weeks following infection. SWAP-specific IgG responses were first detected in the co-infected mice or S. mansoni infected mice at week 4 and 6, respectively. In both groups, SWAP IgG specific responses were similar from week six up the time the experiment was terminated at week 10 (Fig. 3A). When responses to SLA were considered, IgG responses in L.major infected mice or coinfected mice, were not detected until week 10 post infection when some IgG responses were observed (Fig. 3B). For Fig. 3A and 3B, Sera were obtained at regular intervals, pooled and assayed for IgG against either SWAP (A) or SLA (B). Data shown represent means for each group. Solid square, S. mansoni infected group (SM); Solid diamond, L. major infected group (LM); Open triangle, Co-infection of S.mansoni and L. major (SL); Broken lines with long dashes, sera from positive control (CP); Broken line of small dashes, sera from a negative control (CN). Discussion The current study has demonstrated two interesting observations: one, that co-infection of S. mansoni and L. major exacerbated cutaneous lesions, and two, that the co-infection results in less severe S. mansoni induced liver pathology. In an attempt to understand pathogenesis of the co-infection, immune responses to antigens of either parasite were compared in both co-infected and a single parasite infected mice. Apparently, the spleen cell proliferative responses to SWAP were the same in either mice infected with S. mansoni alone or the co-infected animals. Interestingly, some low proliferative responses to SWAP were observed in animals infected with L. major alone, probably a case of shared antigens [8]. Responses to Leishmania antigen were observed in both L. major infected and co-infected groups. However, responses in the co-infected group declined with establishment of the S. mansoni infection. It is possible that there is reduced cellular response to Leishmania antigen with establishment of S. mansoni infection. A low response was observed in the S. mansoni group at week 6, probably, like explained before, a case of shared antigens. Proliferative responses to Con A was not affected in anyway in all the groups indicating that both the single and co-infection did not depress the immune system. As expected, the naïve controls’ spleen and lymph node cells demonstrated background responses [14]. Antigen-specific IgG responses to either S. mansoni or L. major antigen were basically low but positive with similar trend in either L. major only infected or co-infected mice. However responses to S. mansoni were more amplified than responses to L. major. Single infection in S. mansoni stimulates Th2 responses [13]. This would boost the Th2 response, which is also operating at the same time due to L. major infection in the BALB/c [7]. Footpad measurements revealed that by week 10, the co-infected mice had significantly larger nodules as compared to the L. major infected mice (p<0.05). It appears that S. mansoni infection boosted nodule formation process initiated by L. major resulting in a more aggravated and larger nodule formation in the co-infected group. The mice infected with S. mansoni alone had more aggravated liver pathology as compared to the co-infected group. This was demonstrated by: larger lymph nodes, severe liver pathology and high mortality rate at week 10. Unlike the singly infected S. mansoni animals, all the co-infected mice survived to week 10. It appears as if L. major modulated the schistosomiasis-related pathology. It is possible that L. major infection triggered a mechanism leading to direct killing of the schistosome parasites. This in turn could have led to lower numbers of adults laying eggs, and hence reduced pathology. In the co-infected mice, Leishmania seems to modulate S. mansoni infection, while S. mansoni seems to exacerbate L. major infection. Could this divergence be explained by having the two arms of immunity operational at the same time in the coinfected mice? Cellular responses protecting against S. mansoni and high IgG levels leading to exacerbated L. major response? Yoshida [13] challenged S. mansoni-infected mice with and L. major. However, the already existing S. mansoni infection did not affect the outcome of L. major, infection in both susceptible BALB/c and resistant C57BL/6 mice. The difference between their results and the results reported here could be the timing of the infection. In our work, both the S. mansoni and L. major, infections were performed at the same time, while Yoshida [13] infected mice, having an established S. mansoni infection, with L. major. Despite their genetic differences, S. mansoni and L. major, could be sharing some antigens as demonstrated by proliferative responses in each of the single infection groups, to the antigen of the parasite not used to infect that particular group. ConclusionObservations made from this work suggest that coinfections of S.mansoni and L. major result in two different outcomes: one, the co-infected mouse is better protected against S. mansoni pathology, and two, the pathology due to L. major is exacerbated as shown by the significantly larger nodule sizes of coinfected mice. AcknowledgementsWe thank Simon Kiarie, Fred Nyundo, Sammy Kisara, Esther Kagasi and John Machariah for their technical assistance. References
Copyright 2007 - African Forum for Health Sciences The following images related to this document are available:Photo images[jh07011f2b.jpg] [jh07011f2a.jpg] [jh07011f3b.jpg] [jh07011f3a.jpg] [jh07011f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}