 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Health Sciences, Vol. 14, No. 3-4, Jul-Dec, 2007, pp. 187-194 A comparative study of Multiple versus Single infection doses of Schistosoma haematobium in Golden hamsters (Mesocricetus auratus) Vincent K. Njoroge1, Fred Nyundo2, Moses Limo1 and Dorcas S. Yole2 1Egerton University, Department of Biochemistry and
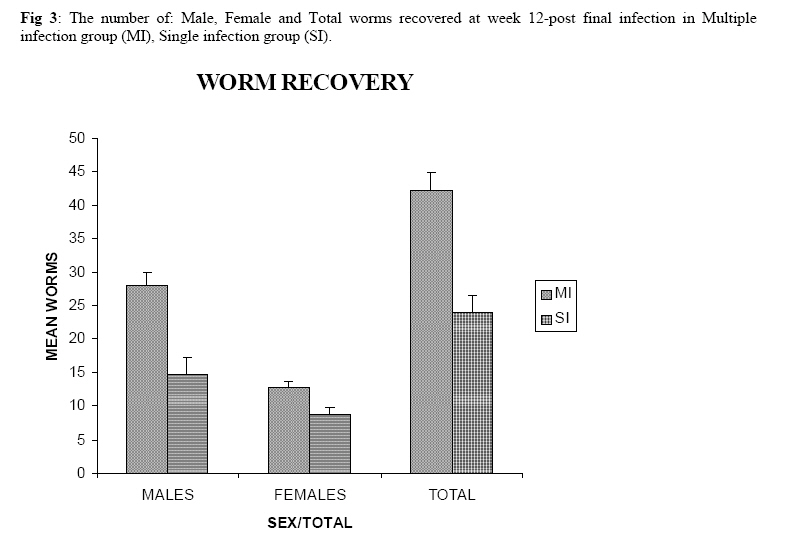
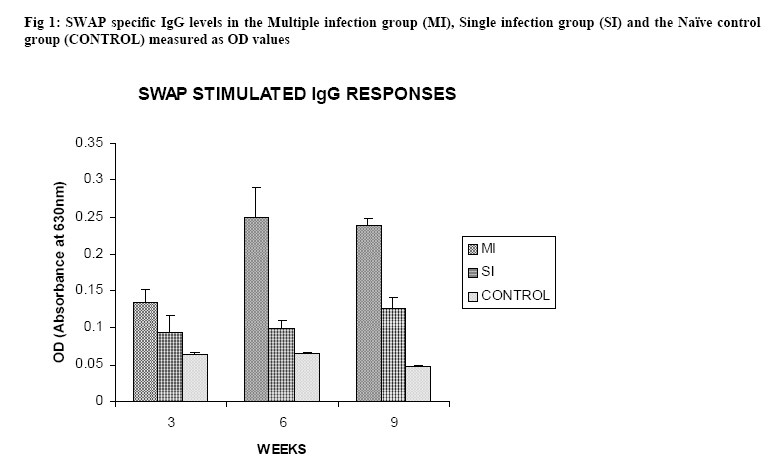
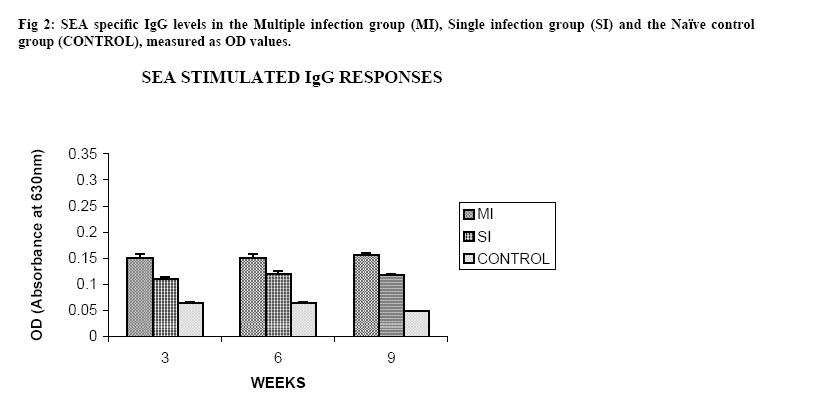
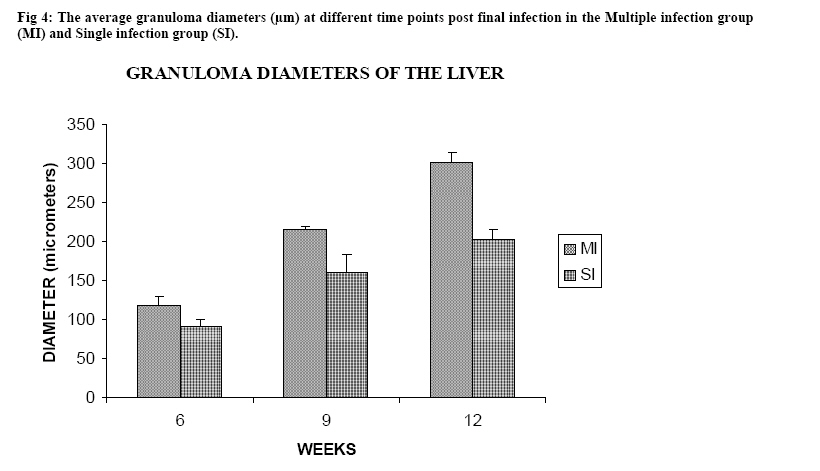
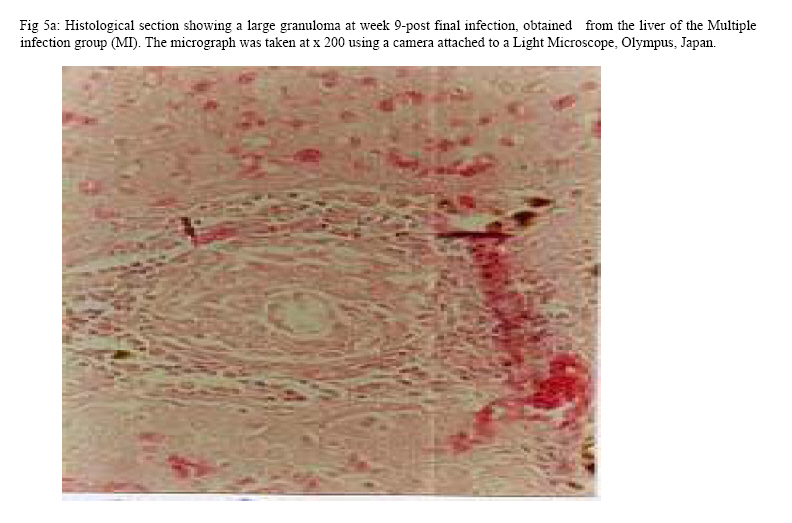
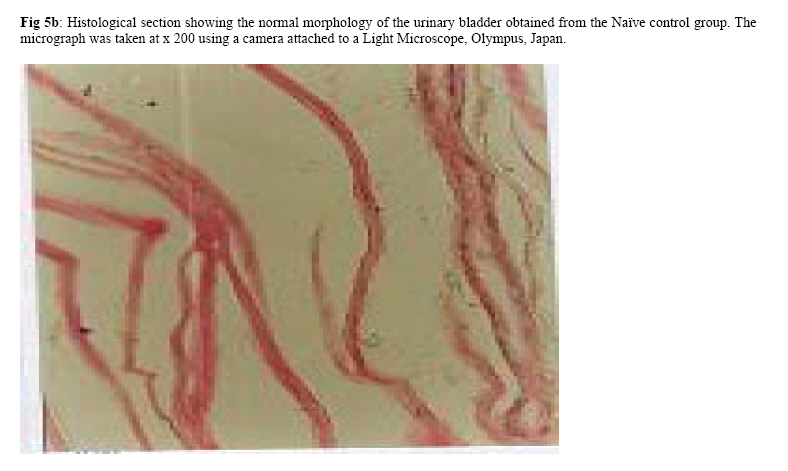
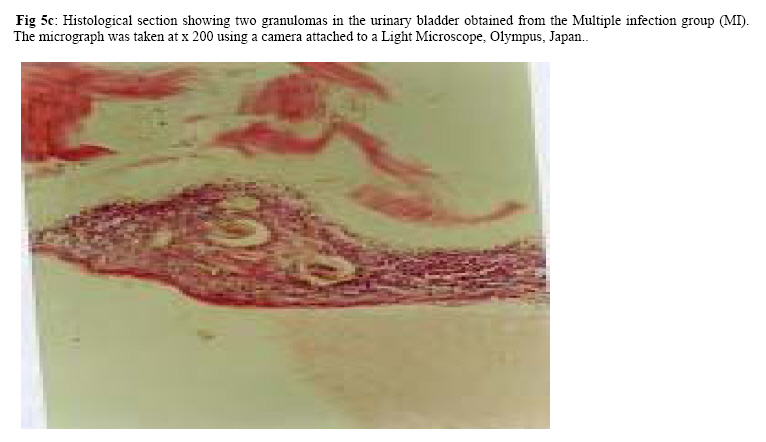
Molecular Biology. P.O Box 536, Njoro, Kenya; Code Number: jh07027 SUMMARY Schistosoma haematobium is widely distributed in Africa. In Kenya, endemic areas include the upper and the lower regions of the Coast Province, Lake Victoria and the Kano plains. Low infection rates over prolonged periods of time characterize schistosome infections of people living in endemic areas. However, the common laboratory practice is to expose the definitive host to a single high dose. In order to utilize the laboratory results appropriately, it is important to know whether or not a large single infection has similar results to multiple small doses. In this study, immune responses, worm burden, gross and histopathological patterns of multiple infection of S. haematobiumium in the Golden hamster (Mesocricetus auratus) were compared with those of single exposure. Multiple infections with low doses of the parasite did not seem to be protective, as suggested by; more worms, worse gross and histopathology in multiple low dose group compared to single high dose group. Most probably there is an antigenic threshold, which needs to be attained before protective mechanisms come into play. Although necessary for protection, there was no direct correlation between IgG levels and degree of protection. Introduction Schistosomiasis is a parasitic disease that ranks second to Malaria in terms of global health importance. Over 200 million people are infected worldwide [1], with 85% of this population living in sub-Saharan Africa. Schistosoma haematobium, the parasite that causes urinary schistosomiasis, is widely distributed in Africa. Seventy million people are infected with S. haematobium with an annual mortality of 150,000 [2]. In Kenya, endemic areas include the upper and the lower regions of the Coast Province, L.Victoria and the Kano plains [3]. Factors that determine the level of morbidity in an exposed population include: thedegree and length of exposure [4], intensity of infection, co-infection [5] and ability to modulate those reactions [4]. Low infection rates over prolonged periods of time characterize schistosome infections of people living in endemic areas. Protective immunity has been shown to be impacted on the Olive baboon (Papio cynocephalus anubis)after multiple exposure to S. mansoni normal cercariae [6]. However, it is not known whether or not a similar scenario occurs in Schistosoma haematobium. In this study, immune responses, worm burden, gross and histopathological patterns of multiple infection of S. haematobium were compared with those of single exposure. Golden hamsters (Mesocricetus auratus) were used as the animal model. Methodology Parasites Bulinus globossus snails were used as intermediate host. The snails were obtained from a dam in Mazeras along the Kenyan Coast. They were exposed to light for 5 consecutive weeks to exclude wild S. haematobium infection. The snails were maintained in the Institute of Primate Research Malacology laboratory at Fort Jesus, Mombasa. Urine from S. haematobium positive patients was obtained from pupils of Kajiwe Primary School, Mazeras. Eggs were recovered by filtration method and thereafter hatched to produce miracidia.The snails were placed individually in each well of a 24-culture plate. Each snail was infected with 3-6 miracidia. The miracia were left for 30 minutes to penetrate. The infected snails were placed in a glass aquarium tank containing 18 l of de-chlorinated water (snail water), sterilized sand and pebbles with some aquatic plants. Three weeks post infection the snails were transferred to a dark room to prevent shedding of cercariae. After five weeks of infection, snails were removed and put under strong light to induce shedding of cercariae. Definitive hosts Golden hamsters, Mesocricetus auratus, bred in the IPR Rodent facility, were dividend into three groups of 20 hamsters each: Multiple infection group (MI), Single infection group (SI) and the Naïve control group (NC). Each group had equal number of males and females. All the hamsters in the Multiple infection (MI) group were anaesthetized using Ketamine (Agrar, Holland) and Xylaxine (Rompun, Bayer Leverkusen, Germany) mixture in the ratio 20:1, at 0.02ml/30g body weight via intraperitoneum. The anaesthetized animals were infected with 50 normal S. haematobium cercariae by abdominal skin exposure using the ring method [7]. Infection with the same dose was repeated for three consecutive weeks therefore, each animal received a total of 200 cercariae. Hamsters in the Single infection (SI) group were anaesthetized as above and were all infected with single dose of 200 normal S. haematobium cercariae during the final infection of the MI Group. Control animals were not infected with S. haematobium. Sampling time points At weeks 3, 6 and 9 post final infection, five hamsters from each of the three groups, MI, SI and NC were examined. Blood was obtained for serum preparation; gross observations were made and tissues (livers & urinogenital system) were obtained for histopathological analysis. At week 12-post final infection, five animals from SI and MI groups were perfused for worm recovery. The experiment was repeated twice for all the groups. Soluble worm antigen (SWAP) and Soluble egg antigen (SEA) preparation The SWAP and SEA antigens were obtained from S. haematobium mature worms and eggs, respectively, using sonication and uitracentrifugation techniques as described by Yole et al., (1996) [8]. Detection of SWAP by immunoglubulins in hamster serum would review presence of adult worm antigens in vivo at the time when the blood was drawn from the hamster, implying presence of adult worms; SEA would demonstrate egg antigens in vivo, hence presence of eggs. However, the different stages of schistosomes share some antigens. Schistosome Specific IgG Enzyme Linked Immunosorbent Assay (IgG-ELISA): IgG is the immunoglobulin used characteristically in testing of schistosome infections. Blood was obtained from the experimental and control groups at weeks 3, 6 and 9 post final infection, using the heart puncture technique and placed in 15 ml centrifuge tubes.Blood was centrifuged at 1000g for 10 minutes and sera retrieved. Immunolon-4 plates (Dynatech, USA) were coated with 50µl of 10µg/ml of SWAP or SEA individually in PBS/0.05% tween-20 buffer (PH 9.6) and incubated at 40C overnight. The plates were blocked with 100µl of 3% bovine serum albumin in PBS/ 0.05% Tween-20 per well and incubated at 37 0C for 1 hour after which they were washed three times with PBS/0.05% Tween-20 using the automated ELISA plate washer. The hamster serum was diluted 1:100 in 1% PBS/0.05% Tween-20 and 50µl of the diluted serum was dispensed into each well. The plates were incubated for 2 hrs at 370C, after which the plates were washed as above. Goat anti-mouse IgG conjugated to hoarse radish peroxidase (Jackson’s Immunoresearch laboratories INC) was diluted 1:2000 in 3% Bovine Serum Albumin (BSA). Fifty microlitres of the diluted conjugate was added into each well. The plates were incubated for 1 hrs at 370C then washed as above. Fifty microlitres of peroxidase substrate per well was added (Substrate used was TMB micro well peroxide substrate from Kirkegaard and Perry labs, USA). After 15 min, the Optical Density (OD) values were generated automatically at an absorbance of 630nm using Maxi Kinetic micro plate reader (Molecular Devices, Palo Alto, CA). Adult worm recovery At week 12-post final infection, five hamsters from each group were perfused according to a modified method of Smithers and Terry 1965 [7]. Adult worm recovery was done according to the method described by Yole et al., 1996 [8]. Gross and Histopathological examination At weeks 6, 9 and 12-post final infection, gross pathology of the liver and the urinogenital system were observed in all the groups. The indices of comparison were; colour, size and presence or absence of granulomas in the liver. Histopathological representative samples of these organs were made and analysed using the method of Farah et al., (1997) [6]. Briefly, tissues were fixed in formalin. Representative samples were cleared in toluene, infiltrated in hot paraffin and embedded on tissue-embedding paraffin wax (Sherwood Medical Co USA). The tissues were sectioned serially at 6 µm using a rotary microtome. The thin tissue sections were stained with hematoxylin and eosin dyes. Slides were mounted and tissues observed under light microscope. Only granulomas containing an ovum in the center were enumerated and measured at 100x magnification using a calibrated ocular micrometer. The averages of the horizontal and vertical diameter were taken to be the diameter of the granuloma. Statistical analysis Worm burden, ELISA data, gross and histopathological results, were analysed using the Students’ two-tailed t test; p<0.05 was regarded as significant. Results Worm burden The mean number of S. haematobium worms recovered in the MI group at week 12-post final infection, was 42.2±2.71 while the SI group had a mean number of 24.0±2.59 of worms. The means of MI and SI groups were significantly different from each other (p<0.05, Students’ t-test; Fig 3). This translated to a worm maturation of 21.1% in the MI group and 12.0% for the SI. In both groups, there were more male than female worms. IgG responses IgG response to SWAP in the MI group were significantly higher than those for SI. (p<0.05, Students’ t test; Fig 1). Peak responses for MI were at week 6-post final infection, while those for SI were at week 9 post final infection. Responses for the Experimental group were higher than in the control group. Similar responses were observed for SEA, with MI having higher responses than SI, throughout the entire sampling period (Fig 2). However, for each of the experimental groups, there was hardly any variation in IgG levels for the three examination points. Gross pathology Granulomas in the MI group were first observed at week 9-post final infection the livers were enlarged. In the SI group the granulomas were not visible and the livers were only slightly enlarged at this time point. By week 12-post final infection, granulomas in the MI group were numerous and the livers were massively enlarged. The SI group livers were enlarged but only a few granulomas were visible. No clear pathology was observed in the urinary bladders in the two groups at these time points. Histopathology At week 6-post final infection, the MI group exhibited many small granulomas on the liver with an average diameter of 118±12.3 µm (Fig 4). Only little cellular infiltration was observed around the portal veins. On the other hand, the SI group had only few granulomas with an average diameter of 91+8.6 µm. At week 9, the MI group showed a significant increase in granuloma diameter as compared to week 6, with an average diameter of 214±6.8 µm. Fibrotic tissues were observed and there was massive cellular infiltration around the portal triad. At this time point the mean granuloma diameter in the SI group was 161.3±21.6 µm and mild cellular infiltration around the portal triad was evident. Peak granuloma size was recorded at week 12 for the two groups. At this time, the MI group had a mean granuloma diameter of 310±13.3 µm. A centrally placed disintegrating egg characterized the majority of granuloma in the MI group; the granulomas had concentric layers of fibrotic tissues surrounding the less conspicuous egg (Fig 5a). SI group granulomas had a mean diameter of 203.0±1.33 µm at week 12. The eggs had not started disintegrating and coalescing granulomas were evident. Throughout the examination points the granuloma diameters in the MI group remained significantly greater than in the SI group (p<0.05, Students’ t-test). At weeks 6 and 9 post final infection, histopathological examination revealed normal urinary bladder morphology same as the one observed in the naïve control hamsters (Fig 5b). Granulomas were observed in the urinary bladder at week 12 in the MI group. At this time point the MI group had a mild epithelium congestion of the urinary bladder with massive cellular infiltration along the muscle layers and the bladder veins. Few large granulomas were also evident (Fig 5c). The SI group had mild cellular infiltration along the veins and no granulomas were observed. Discussion In this study, multiple low doses of infection by Schistosoma haematobium, as it occurs in human beings living in endemic areas was studied in an experimental set up using the Golden hamster (Mesocricetus auratus). This was compared with a single high dose infection. Single high dose infection is the method used routinely in research laboratory to study schistosome infections. The immunological responses elicited by the two modes of infection were compared. The immunological index of the test was stage specific Immunoglobulin G (IgG) levels at different time points post schistosome infection. The protective capacity of the responses was accessed by comparing worm burdens, gross pathology and histopathological effects on the liver and the urinary bladder at different time points after infection. IgG in schistosome infection participates in the killing of miracidium inside the egg. The killing is mediated by eosinophils, monocytes and platelets [9]. At week 3 post-final infection, the level of IgG was already high in the two groups, pointing to the importance of IgG during an infection of S. haematobium in hamsters. Throughout the experiment, both SWAP and SEA specific IgG levels were higher in the Multiple infection hamsters compared to the Single infection group of hamsters. As the weeks progressed SWAP specific IgG levels rose significantly in the two groups indicating an increase in stage specific antigens as the parasite matured. SEA specific IgG was high throughout the experiment, suggesting cross reactivity with other stage-specific antigens, for example larval antigens at wk 3 post-infection in the Single infection. This kind of reaction was also observed by Yole et al 1996 [8], were they reported similar responses in baboons.. The Multiple infection group had a higher worm burden and consequently higher worm maturation compared to the Single infection group. It also had worse gross pathology and histopathology examination outcomes. A higher number of worms could have translated to higher oviposition and thus the worse gross pathology and histopathology observed in the Multiple infection group compared to the Single infection group. This suggests that, multiple infection of hamsters with small numbers of S. haematobium cercariae does not trigger a protective immunological response. The probability is that there is an antigenic threshold, which needs to be attained for initiation of protective mechanisms in vivo. Our results contradict observation made by Farah et al. (1997) [6], who obtained a lower number of Schistosoma mansoni worms in baboon receiving small multiple doses, as compared to those receiving a single high dose. This could be attributed to the use of different species of the parasites and different definitive hosts. The Multiple infection group had a higher worm burden and higher levels of IgG antibody compared to the Single infection group, pointing to the fact that, although IgG is required for protection in schistosome infection, the levels may not be proportional to the degree of protection. Most of the IgG could be produced against schistosome antigens not important for protective immune mechanisms, for example, internal antigens. Alternatively, it could be due to the fact worms have devised an immune evasion mechanism against the IgG. This phenomenon compares well with what was observed by Mduluza et a., 2001 [10] who reported presence of suppressive factors in serum of individuals who had long durations of infection. Conclusion A large single dose of Schsistosoma haematobium seems to elicit a better protective mechanism in Golden hamsters, against the parasite compared to multiple small doses of the parasite. Although necessary for protection, there is no direct correlation between IgG levels and worm reduction in our experiment. Acknowledgement We wish to thank Sammy Kisara, Shadrack Rakamba and Mary Nzuve for their technical support during this study. References
© Copyright 2007 - African Forum for Health Sciences The following images related to this document are available:Photo images[jh07027f1.jpg] [jh07027f5b.jpg] [jh07027f5c.jpg] [jh07027f3.jpg] [jh07027f4.jpg] [jh07027f2.jpg] [jh07027f5a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}