 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Postgraduate Medicine, Vol. 48, Issue 3, 2002 pp. 186-190 Lipid Peroxidation and Antioxidant Enzymes in Male Infertility Dandekar SP, Nadkarni GD*, Kulkarni VS, Punekar S** Departments of Biochemistry and Urology,** Seth G. S. Medical College, and
Radiation Medicine Centre (BARC),* Tata Memorial Hospital, Parel, Mumbai - 400
012, India. Code Number: jp02064 Abstract: Background and Aim: Mammalian spermatozoa are rich in polyunsaturated fatty acids and are very susceptible to attack by reactive oxygen species (ROS) and membrane lipid peroxide ion. Normally a balance is maintained between the amount of ROS produced and that scavenged. Cellular damage arises when this equilibrium is disturbed. A shift in the levels of ROS towards pro-oxidants in semen and vaginal secretions can induce an oxidative stress on spermatozoa. The aim was to study lipid peroxidation and antioxidant enzymes such as catalase, glutathione peroxidase and superoxide dismutase (SOD) and to correlate the same, with the "water test", in male infertility. Settings: Experimental study. Subjects and Methods: Ejaculates from a total of 83 infertile and fertile healthy individuals were obtained. Lipid peroxidation and antioxidant enzyme levels were studied and correlated with water test. Results: The results indicate that (i) the antioxidant enzyme catalase showed no significant changes in the various pathological samples, (ii) antioxidant enzymes SOD and glutathione peroxidase correlate positively with asthenozoospermic samples and (iii) the degree of lipid peroxidation also correlates positively with the poorly swollen sperm tails. The increase in SOD and glutathione peroxidase values, in the pathological cases represents an attempt made to overcome the reactive oxygen species. Conclusion: Water test could be used as a preliminary marker test for sperm tail damage by reactive oxygen species, since it correlates very well with lipid peroxidation and antioxidant enzymes. (J Postgrad Med 2002;48:186-190) Key Words: Male infertility, human semen, lipid peroxidation, glutathione peroxidase, superoxide dismutase, catalase, water test. Defective sperm function is the most prevalent cause of male infertility and is difficult to treat.1 Inflammation of the male accessory sex organs resulting in conditions like leucocyto-spermia has been implicated as an important cause of defective sperm motility and function.2 Human spermatozoa represent a growing list of cell types that exhibit a capacity to generate reactive oxygen species (ROS). ROS are important mediators of sperm function.3 Mammalian spermatozoa membranes are rich in unsaturated fatty acids, which are sensitive to oxygen induced damage mediated by lipid peroxidation (LPO).4 Antioxidants, in general are compounds which dispose, scavenge and suppress the formation of ROS or oppose their actions. Of the antioxidants studied, the enzymes, glutathione peroxidase, superoxide dismutase and catalase play significant roles.5 Various tests have also been developed to assess the functional and morphological integrity of the sperm membrane. A common method to evaluate sperm membrane using distilled water, as by Lomeo and Giambersio,6 uses distilled water as a medium of strong hypotonicity leading to swelling. On the basis of the swelling of the spermatozoa tail, categories are differentiated as >80% swelling, taken as the control group showing intact sperm membrane, 61-80% non-swollen tails, 20-60% non-swollen tails and <20% non-swollen tails. Two major questions addressed in this study are to correlate the extent of lipid peroxidation with the "water test" and to correlate the semen samples subdivided according to the classical spermiogram with the release of the different antioxidant enzymes.
Material and Methods Approval for the study was taken from the institutional ethics committe and written, informed consent was taken from all participating subjects. The inclusion criteria for the study were as follows - Age between 25 and 45 years and semen samples devoid of leucocytes7. All samples were obtained from the department of urology, men with proven fertility were taken as controls. Semen samples were collected and classified according to WHO criteria.8 They were divided into four categories: normospermia (> 40 x 106 sperm/ml, 60% motility), severe oligospermia (< 15 x 106 sperm/ml, 20% motility), mild oligospermia (> 15 x 106 sperm/ml, > 40% motility), azoospermia (no sperms seen) and asthenozoospermia (< 10 x 106 sperm/ml, ~ 20% motility). Sperm preparation After 30 minutes liquefaction at room temperature, spermatozoa from all samples were separated from seminal plasma by centrifugation at 500 X g for 10 minutes. Homogenising buffer was added to the separated fractions. Homogenising buffer consisted of mannitol, sucrose, EDTA mixed together with the pH adjusted to 7.4. The samples were hand homogenised and were subsequently centrifuged for 10 minutes at 3000 rpm in a refrigerated centrifuge. Triton X-100 (0.1%) was added to the pellets obtained and the samples were centrifuged again at 8000 rpm for half an hour in a refrigerated centrifuge. This concentration of Triton X-100 does not affect enzyme levels. The supernatant was used for enzymatic measurements. All samples were screened for leucocyte contamination. Only samples devoid of contamination were chosen as leucocytes have been implicated in the production of ROS.9 Lipid Peroxidation The amount of malondialdehyde (MDA) produced was used as an index of lipid peroxidation. This was determined by the thiobarbituric acid (TBA) assay of Uchiyama and Mihara.10 500 ml of homogenate was added to 3.0 ml of 1% phosphoric acid, 1.0 ml of 0.6 % TBA, and 0.15 ml of 0.2% butylated hydroxytoluene ( BHT ) in 95% methanol. The samples were heated in a boiling water bath for 45 minutes, cooled and 4.0 ml of butanol was added. The butanol phase was separated by centrifugation at 3000 rpm for 10 minutes and absorbance was measured at 532 nm. All values were expressed as nmoles MDA present in it and finally expressed as nmoles MDA/mg of protein. Determination of Superoxide Dismutase (SOD) activity SOD was measured according to the method of Marklund and Marklund.11 Superoxide anion radical is involved in the auto oxidation of pyrogallol. At alkaline pH, SOD dismutates superoxide thereby inhibiting the auto oxidation of pyrogallol. The stock pyrogallol solution contained in 1.0 ml of 0.5 mol/L HCl was stored at 400C. The assay mixture contained 500 ml of buffer, 50 ml of sample, 400 ml of deionized water in a cuvette. The blank sample contained 500 ml of assay buffer and 450 ml of water. The absorbance of each sample was measured at 420 nm before addition of pyrogallol. The increase in absorbance was measured at 30 seconds interval upto 3 minutes. SOD activity was expressed as U/mg of protein. Determination of Glutathione Peroxidase (GSH.Px) activity The activity of glutathione peroxidase was analysed by the method of Hafeman et al.12 GSH. Px degrades tertiary butyl hydroperoxide (t-BOOH) in the presence of GSH thereby depleting it. The remaining GSH is then measured by using 5,5' dithiobis - (2 - nitrobenzoic acid) (DTNB). The reaction mixture contained 1.0 ml of 2mM GSH of 0.4 M phosphate buffer (pH 7.0). It also contained 4 x 10-4 M EDTA, 0.5 % of 0.01 M sodium azide, 250 ml of sample was used and water was added to bring the total volume to 4.0 ml. After 5 minutes of incubation at 370 C, 1.0 ml of prewarmed 1.25 mM t-BOOH was added and incubated further for 3 minutes. At the end of 3 minutes, 1.0 ml of reduction mixture was removed and added to 4.0 ml of meta phosphoric acid precipitating solution which was centrifuged for 10 minutes at 3000 to 5000 rpm to obtain a protein free filtrate. The GSH in the protein free filtrate was then determined by mixing 2.0 ml of filtrate with 2.0 ml of 0.4 M Na2HPO4 and 1.0 ml of DTNB reagent. Absorbance of this solution was recorded at 412 nm. A blank was carried through out the incubation simultaneously with the samples, since non-enzymatic GSH oxidation by t-BOOH occurs during incubation. One unit of GSH. Px enzyme activity was defined as the decrease in log [GSH] by 0.001 per minute after the decrease in log[GSH] per minute of the non enzymatic reaction was subtracted. GSH. Px activity was then expressed in U/mg of protein. Determination of Catalase activity Catalase activity was estimated by the method of Aebi.13 It can degrade hydrogen peroxide which can be measured directly by the decrease in the absorbance at 240 nm. The hydrogen peroxide was diluted with phosphate buffer pH 7.0 and its initial optical density was adjusted between 0.5 to 0.6 absorbance unit at 240 nm. The decrease in the absorbance was measured. One unit of catalase activity was defined as the amount of catalase which absorbed in 30 sec at 250C. The catalase activity was then calculated from the change in absorbance and finally expressed as U/mg of protein. Determination of hypoosmotic swelling The "water test" was carried out using the method of Lomeo and Giambersio.6 The test was measured by mixing 10 ml of semen with 40 ml distilled water on a clean glass-slide. The mixture was covered with a thin cover slip and incubated for 5 minutes at 370C, before it was examined with a microscope at x 400 magnification. At least 200 sperms were examined and the percentage of sperm that showed swollen tails was calculated (number of swollen sperms x 100, divided by total number of sperm counted). All data is presented as mean±standard deviation, analysed by one-way analysis of variance (ANOVA), followed by the students `t' test.
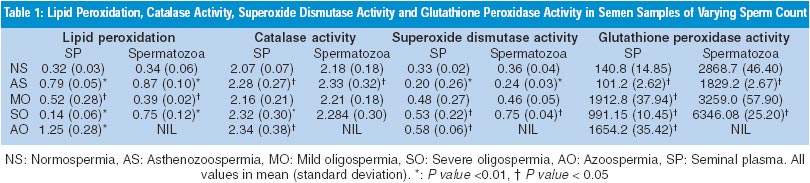
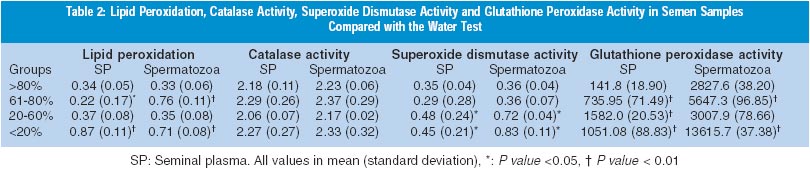
Results A total of 83 samples were studied. Of these, 15 were normospermic, 15 asthenozospermic, 10 mild oligospermic, 33 severe oligospermic and 10 azoospermic. The samples, other than azoospermic, were also categorised on the basis of percentage of swelling of tails; samples with >80% swelling (n=15) were taken as controls and rest of the groups were considered as `non-swelling' (20 with 61-80%, 15 with 20-60% and 20 with less than <20% swelling). Three samples could not be categorised on the basis of swelling of tails due to technical problems. Lipid peroxidation was compared with varying sperm count in seminal plasma and spermatozoa (Table 1). The poorly swollen sperm (<20%) showed highly significant lipid peroxidation. The 20-60% poorly swollen tails did not show significant lipid peroxidation (Table 2). Catalase activity showed no correlation with varying sperm count or with water test. Superoxide dismutase activity when compared with varying sperm count showed significantly increased levels in severe oligospermia. Superoxide dismutase activity showed a positive correlation when compared with the water test. Seminal glutathione peroxidase correlated positively with the sperm count. Similar to the results from seminal plasma, the enzyme activities from extracted spermatozoa did not differ significantly in various pathological ejaculates. Lipid peroxidation was found to be significantly increased in asthenozoospermics (P <0.01), SOD levels showed significant increase (P <0.01) whereas glutathione peroxidase showed a significance of P <0.05. Glutathione peroxidase activity when compared with the water test showed significantly high levels in all the non-swelling categories except for 20-60% poorly swollen category.
Discussion ROS may cause a defect in sperm function through lipid peroxidation.14 This is because spermatozoa, unlike other cells, are unique in structure, function and susceptible to damage by LPO. Therefore, lipid peroxidation and levels of antioxidants have been implicated in disturbances of sperm function. ROS is produced by a variety of semen components including immotile or morphologically abnormal sperma-tozoa, leukocytes, and morphologically normal but functionally abnormal spermatozoa.15 The SOD level in spermatozoa is positively correlated with sperm motility.16 Also, catalase, which prevents ROS damage,5 has been found in both human spermatozoa and seminal plasma. Similarly, a selenium-containing glutathione enzyme scavenging system exists in the sperm of several mammalian species, including human beings.17 This system may act directly as an antioxidant and an inhibitor of LPO. A high GSH/GSSG ratio will help spermatozoa to combat oxidative insults. Normally, a balance is maintained between the amount of ROS produced (pro-oxidants) and that scavenged by a cell (antioxidant). Cellular damage arises when this equilibrium is disturbed, especially when the cellular scavenging systems cannot eliminate the increase in ROS and correlate with idiopathic infertility. The presence of catalase-like activity observed in ROS-producing semen samples may be a compensatory mechanism that produces more low molecular weight H2O2 scavengers. From the study it seems that lipid peroxidation of human spermatozoa may cause loss of motility. Superoxide dismutase and glutathione peroxidase may come into play in order to inhibit the lipid peroxidation. Perhaps, the significant increase in the pathological conditions helps to overcome the lipid peroxidation (Tables 1 and 2). Infertile males have a failure of expression of phospholipid, hydroperoxide, glutathione peroxidase in the spermatozoa.18 Most of these men had defective sperm count and motility. This is in agreement with our results, in which astheno-zoospermic patients showed decreased SOD and glutathione peroxidase levels. It is possible that membrane fragility causes abnormalities, affecting ion exchanges essential for maintaining normal motility. Also, peroxidation may result in hyperpermeability. The discrepancy in the levels of lipid peroxidation in azoospermia, in spite of the lack of sperm and leucocytes, may be due to the lower antioxidant levels found in infertile men. That the lipid peroxide in the seminal plasma does not arise from the spermatozoa, since it was detected from the patients with azoospermia has been shown.19 Surprisingly, catalase levels did not show any significant changes in the pathological groups. It appears that the scavenging systems are defective or inadequate and perhaps the ROS generated significantly decrease the fertilising potential of the sperm. High SOD and glutathione peroxidase activities were observed in 20-60% non-swelling and <20% non-swelling samples. This correlated positively with the degree of lipid peroxidation seen in the samples. Various tests have been developed to assess the functional and morphological integrity of the sperm membrane. The ability of sperm to swell when subjected to a hypo-osmotic solution containing fructose and sodium citrate has been studied. 20 On similar lines, use of distilled water as a medium of strong hypotonicity was studied.21 This was found to be very convenient and economical. The types of swelling have been correlated with lipid peroxidation. The results indicate that "water test" correlates well with lipid peroxidation and with antioxidant enzymes glutathione peroxidase and superoxide dismutase.
Support Mr. V. Kulkarni has received a fellowship from Lady Tata Memorial Trust.
Part of the data was presented at the International Conference on Reproductive Health, 15-19 March 1998, Mumbai.
References
Expert's Comments Mammalian spermatozoa are rich in polyunsaturated fatty acids and are very susceptible to attack by free radicals called reactive oxygen species (ROS) resulting in increased membrane lipid peroxidation. This study correlates the extent of lipid peroxidation in spermatozoa in infertile men and evaluates role of antioxidant enzymes present in the seminal plasma. The authors evaluated lipid peroxidation and antioxidant enzymes such as catalase, glutathione peroxidase and superoxide dismutase (SOD) in semen samples and correlated these with a "water test" also commonly known as hypo-osmotic swelling test "(HOST)" in order to evaluate sperm viability and/or integrity. The authors have confirmed some of the previously published findings. Their results indicate that (i) the antioxidant enzyme catalase showed no significant changes in the various pathological samples, (ii) antioxidant enzymes SOD and glutathione peroxidase correlate positively with asthenozoospermic samples and (iii) the degree of lipid peroxidation also correlates positively with the poorly swollen sperm tails. The increase in SOD and glutathione peroxidase values, in the pathological cases represents an attempt made to overcome the reactive oxygen species. Although the authors conclude that this "water test" could be used as a preliminary marker test for sperm tail damage by reactive oxygen species, since it correlates very well with lipid peroxidation and antioxidant enzymes, they need to explore this by utilizing much better tools and procedures. Of course the effect on sperm fertilising ability is the most important issue. In this study, the groups are classified as normospermia, "severe" and "mild" oligospermia, azoospermia, asthenozoospermia. Although this information is very important and critical for correlations and statistical interpretation of the data, there are overlapping gray zones. Another critical group to evaluate is asymptomatic idiopathic infertility group characterized by leukocytospermia. Once activated, the WBC present in semen samples of such men are the main source of ROS and very damaging to spermatozoa. Future studies should focus on these gray zones and other critical areas. In a normal cell, there is a balance between the amount of ROS produced under various metabolic conditions and the ROS scavenging capacity of antioxidant enzymes. Whenever there is a shift in this balance towards increased ROS that results in oxidative stress, the cellular metabolism changes. In semen and vaginal secretions, increased ROS levels are likely to induce significant oxidative stress on spermatozoa that impairs their motion and function. Oxidative stress in infertility has recently attracted a great attention because of potential toxic effects of free radicals to spermatozoa in both male and female reproductive tract. Recent studies have demonstrated spermatozoa DNA damaging effects of ROS. This area has much greater implications considering the impact on germ cells and transfer of genetic information to progeny. Although presence of a number of antioxidants will play a great role in preventing such damage to spermatozoa, there is lack of knowledge on what antioxidants and how much of these supplements are beneficial.
Sikka SC Department of Urology and Andrology Clinical Laboratory, This article is also available in full-text from http://www.jpgmonline.com/ © Copyright 2002 - Journal of Postgraduate Medicine The following images related to this document are available:Photo images[jp02064t2.jpg] [jp02064t1.jpg] |
| |||||||||

{kind=link}
{kind=link}