 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Postgraduate Medicine, Vol. 50, No. 1, January-March, 2004, pp. 17-20 Original Article Associations of antifolate resistance in vitro and point mutations in dihydrofolate reductase and dihydropteroate synthetase genes of Plasmodium falciparum Biswas S Malaria Research Centre (Indian Council of Medical
Research), 22 Sham Nath Marg, Delhi - 110054
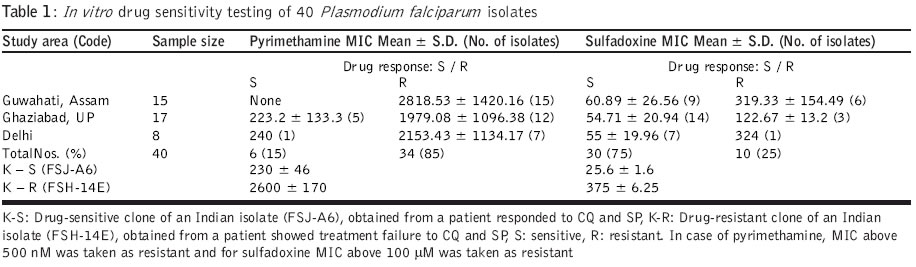
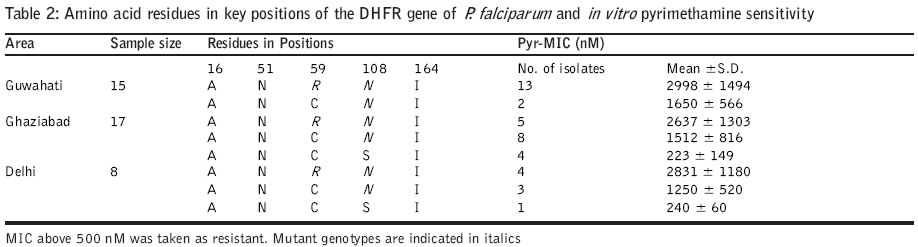
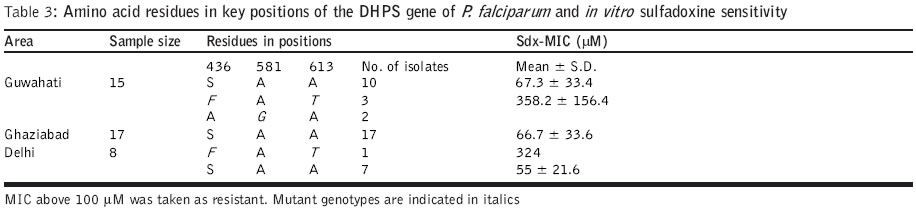
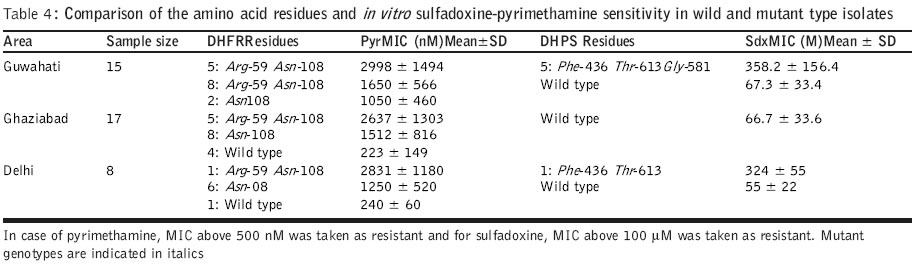
Code Number: jp04006 Abstract Background: Antifolate antimalarials like sulfadoxine-pyrimethamine are used as second-line treatment for Plasmodium falciparum malaria patients who fail to respond to chloroquine. The efficacy of the sulfa-pyrimethamine combination in the treatment is also compromised by the development of resistance in the parasite. Resistance to these drugs has been shown to encode with point mutations in dihydrofolate reductase and dihydropteroate synthetase genes. Settings: An experimental study. Maerial and methods: Forty clinical isolates collected from different geographical locations in India were used to assess the relationships between resistance to sulfadoxine-pyrimethamine (SP) and mutations in P. falciparum dihydrofolate reductase (DHFR) and dihydropteroate synthetase (DHPS). In vitro drug susceptibility and mutation-specific polymerase chain reaction (PCR) assays were also done. Results: It was observed that a number of isolates possessed mutant genotypes and showed low sensitivity to SP in vitro. Of the 40 clinical isolates studied, 87.5% had DHFR and 15% had DHPS gene mutations. As observed from PCR results, 55( (22/40) presented double mutation of DHFR Arg-59 and Asn-108 and 32.5 % (13/40) had single mutant type allele of Asn-108. Of the 40 isolates, 10 % (4/40) presented doubly mutated forms of DHPS Phe-436 and Thr-613 and single mutant type allele Gly-581 was detected in 5 % (2/40) isolates. Parasites carrying double or single mutant forms of DHFR/DHPS showed elevated minimum inhibitory concentration (MIC) values of both pyrimethamine (760–6754 nM; r=0.69) and sulfadoxine (108 – 540 mM; r=0.87) when compared to sensitive and resistant strains. Conclusion: Though there was a correlation between molecular techniques and in vitro drug sensitivity profiles, the relevance of these findings to the clinical efficacy of SP combination drugs needs to be established by controlled clinical trials.Among the Southeast Asian countries, India alone contributes more than 80% cases of malaria and P. falciparum accounts for 35-40% of these cases.[1] Due to increasing chloroquine (CQ) resistance, the antifolate (sulfadoxine-pyrimethamine) drugs are gaining importance in the treatment of uncomplicated falciparum malaria.[1],[2],[3] Sulfadoxine-pyrimethamine (SP) is used in synergistic combination.[4] The compounds interfere with the synthesis of parasite DNA from guanosine triphosphate (GTP) and thus act as folate pathway blockers. Pyrimethamine binds and inhibits dihydrofolate reductase (DHFR) and sulfa drugs act on dihydropteroate synthetase (DHPS).[5],[6] A single point mutation causing a Ser ®Asn change at codon 108 is linked to pyrimethamine resistance. The addition of Asn ®Ile-51 and / or Cys ®Arg-59 mutations confers higher level of pyrimethamine resistance. Ser ®Thr-108 coupled with Ala ®Val-16 is associated with resistance to cycloguanil (the active metabolite of proguanil, a DHFR inhibitor). Ile ®Leu-164 combined with Asn-108 plus Ile-51 and / or Arg-59 confers high level of resistance to both pyrimethamine and cycloguanil.[5],[7],[8] P. falciparum strains with variable levels of sensitivities to sulfadoxine demonstrated sequence variation in DHPS. Sulfadoxine resistant strains showed mutations at Ser-436 to Phe-436; Ala-437 to Gly-437; Ala-613 to Ser or Thr-613 as well as a single point mutation at Ala-581 to Gly-581. Additional mutation at Lys-540 to Glu-540 was also reported in isolates from Thailand, Bolivia, Kenya and Tanzania.[8],[9],[10],[11] For monitoring drug sensitivity patterns, it is essential to have a simple in vitro test for field evaluation of susceptibility of parasite isolates to the drug. An in vitro micro-culture system for the evaluation of synergy between pyrimethamine and sulfadoxine has been developed.[4],[11] I herewith report studies on field isolates of P. falciparum collected from different geographical locations of India to compare their in vitro drug sensitivity status against sulfadoxine and pyrimethamine and the prevalence of point mutations in the DHFR and DHPS genes for determining association between the two methods. Material and Methods Forty patients with P. falciparum infection who were 1 year or older were enrolled into the study. (3: 1-5 yr; 8: 5-14 yr; 29: 15 years and above) during the transmission season in the years 2000 and 2001. Study subjects were inhabitants of northern and northeastern parts of India. P. falciparum isolates were obtained from local malaria clinics of Delhi (n=8) and a village of Loni (n=17) P.H.C., Ghaziabad, UP in north India and from a local hospital of Guwahati (n=15), Assam of the northeastern zone. Information on the occurrence and duration of fever and the use of antimalarials was recorded. P. falciparum smear positive patients having 1000 or more asexual parasites per microlitre blood were included in the study. 200-300 ml of blood was collected from each patient by finger prick into sterile heparinized capillary tubes. Patients diagnosed with malaria were treated with antimalarials as per recommendation. The study was initiated after approval of the institutional ethics committee and after obtaining written informed consent.Parasite culture and in vitro drug sensitivity assays: P. falciparum isolates were cultured by the established method.[12] The growth medium used for culture and drug sensitivity testing was RPMI 1640 devoid of added p-aminobenzoic acid or folic acid (GIBCO, UK), supplemented with 25mM HEPES buffer and sodium bicarbonate and enriched with 10% human AB+ serum (obtained from a blood donor belonging to malaria non-endemic area). Drug sensitivity assays[4],[13],[14] for pyrimethamine (Pyr) and sulfadoxine (Sdx) were done for all 40 isolates. Flat-bottomed 96 well tissue culture plates were dosed separately with various concentrations of Pyr (0.5-50,000 nM) and Sdx (0.4-5000 mM). An aliquot of 100 mL of infected erythrocyte suspension was added in the control and drug-dosed wells. The plates were kept in a candle jar and incubated at 370C for 30 hours. After incubation, the supernatant was aspirated, the pellet was mixed and thick / thin smears were prepared for microscopic examination. As SP acts on late stages, schizonts having 8 or more nuclei were counted/ 200 asexual parasites. The assays were considered successful if 10% or more of the parasites in the control wells developed into schizonts with 8 or more nuclei. The minimum inhibitory concentration (MIC), the lowest drug concentration in which no such schizonts were observed, was determined using probit analysis for individual isolates. The parasite genomic DNA was extracted from parasitized erythrocyte pellet by proteinase K treatment followed by phenol:chloroform extraction, ethanol precipitation and reconstitution of extracted DNA in sterile triple distilled water.[15] The PCR primers spanning a predicted product of 708 / 720 nucleotides were selected to amplify the entire DHFR gene.[5],[16] The amplified DHFR gene fragment (708 bp) was digested separately with Alu I, Bsr I, ScrF I and Nla III restriction endonucleases to detect point mutations. Digested products were immediately separated on 1.5% agarose gel, then stained and visualized under UV. Digestion with Alu I (codon Ser-108), Bsr I (codon Asn-108) and ScrF I (codon Thr-108) produces 2 bands of 386 and 322 bp. Nla III digestion of the wild type (codon Ala-16) produces 3 fragments of 568, 93 and 47 bp, whereas mutant type (codon Val-16) produces 2 bands of 568 and 140 bp. Along with test DNA samples, known controls of both wild (3D7) and mutant (HB3 and FCR3) DHFR genotypes were also tested. Nested mutation-specific PCR was done to analyse the mutations at 3 codon sites of the DHFR gene using published methodology.[8],[16],[17] Amplified DHFR domain (720 bp) from the first round of PCR was used in the mutation-specific second round of PCR to assess the mutation at 51 (563 bp), 59 (190 bp) and 164 (505 bp) codon sites. The PCR assay conditions were followed as previously described and the amplified products were analysed on 1.5% agarose gel containing ethidium bromide. DHPS gene amplification, mutation-specific PCR: The sequences of the primers and procedure were followed as per published methodology.[8],[18] Primary PCR amplified the DHPS gene using two flanking primers, which yielded a product of 1.15kb. Amplified DHPS domain from the first round of PCR was used in the second round nested PCR by using two flanking and seven mutation-specific oligonucleotide primers to detect point mutations at codon sites 436 (876 bp), 581 (758 bp) and 613 (852 or 849 bp). Mutations at 437 and 540 codon sites could not be done due to the non-availability of the reagents. Known sulfadoxine sensitive (3D7) and resistant (V1/S) clones were taken as control. PCR amplified products were electrophoresed, stained and visualised. Statistical analysis: The differential responses of parasite isolates to Pyr and Sdx were determined by the method of one-way ANOVA. The prevalence of the DHFR-DHPS genes mutation and drug sensitivity was compared by chi-square(c2) test and linear regression analysis was done between Pyr-Sdx sensitivity and prevalence for each mutation. The results were considered significant at the 5% level. Results The Pyr and Sdx susceptibilities of 40 field isolates and 2 reference lines of P. falciparum are shown in [Table - 1]>. The susceptibility patterns of the isolates fell into two significantly different groups (P<0.01) for both drugs when compared with reference sensitive and resistant lines. In case of Pyr, MIC values <500 nM and >500 nM were observed in isolates with wild and mutant genotypes, respectively, whereas in case of Sdx, MIC values less than and above 100 mM were observed in known sensitive and resistant lines. From overall in vitro drug susceptibility profiles, it was observed that of the 40 isolates, 6 (15%) were Pyr sensitive whereas 30 (75%) isolates were Sdx sensitive [Table - 1]>. The MIC values of Pyr-sensitive and Pyr-resistant groups were 118-480 nM and 760-6754 nM, respectively, and that of the Sdx-sensitive and Sdx-resistant groups were 20-98 mM and 108-540 mM, respectively. All 15 isolates from Guwahati showed high MIC values to Pyr [Table - 1]>. In case of Sdx, only 6 isolates showed high MIC values. Of the 17 isolates from Ghaziabad, 12 were resistant to Pyr while 3 were found resistant to Sdx. Among the 8 Delhi isolates, 7 were resistant to Pyr whereas only 1 was found resistant to Sdx. The comparative data of the amino acid residues in key positions of the DHFR and DHPS genes and in vitro Pyr / Sdx sensitivity are given in [Table - 2]>, [Table - 3]> and [Table - 4]>. The results of this study confirm the presence of Pyr-resistant isolates and a few Sdx-resistant isolates among smear positive patients and they are in agreement with in vitro drug sensitivity results. Of the 40 clinical isolates studied, in case of DHFR, 55% (22/40) presented double-mutated forms of Arg-59R and Asn-108; single mutant type allele Asn-108 was detected in 32.5% (13/40) isolates. Mutations at 16, 51 and 164 positions were not observed in any of the isolates. None of these isolates showed paired mutation at position 108 and 16, associated with resistance to cycloguanil and low response to Pyr [Table - 2]>. In case of point mutations in the DHPS gene, 10% (4/40) presented double-mutated forms of Phe-436 and Thr-613, single mutant type allele Gly-581 was detected in 5% (2/40) isolates. None of these isolates showed single mutation at position 436 and 613 [Table - 3]>. Isolates carrying double or single mutations in the DHFR gene showed MIC values above 500 nM (r=0.69; P<0.001), whereas parasites without any point mutation were sensitive to Pyr in vitro. Similarly, isolates carrying double or single mutation in the DHPS gene showed MIC values above 100 mM (r=0.87; P<0.05), whereas parasites without any point mutation were sensitive to Sdx in vitro [Table - 4]>. Discussion Despite the understanding of the molecular basis of the resistance of P. falciparum to antifolate drugs, the determinants of treatment failure with the combination of SP are still under investigation. The work was done to establish a basis for determining the drug sensitivity status and the prevalence of mutation in the field isolates and their possible role in drug-resistant malaria. The present work on DHFR and DHPS mutations and in vitro drug susceptibility in the light of SP treatment reveals the utility of these simple, relatively rapid methods and their application in surveillance programmes to detect drug-resistance mutants.[8] The parasite genotypes linked to pyrimethamine resistance are highly frequent in various parasite populations around the world.[15] In this study also, Pyr-resistant parasites were detected in more than 85% of the samples. The drug-induced selection of Pyr-resistant genotypes is very efficient as reported in earlier studies.[9] The association of the phenotypic resistance of the parasite and the genotype is linked to a common mutation of Asn at position 108 of DHFR.[7],[15] Mutation at site 59 along with 108, contributes resistance against higher levels of the drug as observed in the drug sensitivity profiles in vitro. The antifolate resistance has also been linked with paired mutations of DHFR Val-16 and Thr-108.[14] The prevalence of mutant forms of DHPS also varies among countries; 3% in Cameroon and Kenya, 4.5% in India, almost 30-34% in Tanzania and Thailand, 94% in Mali and 100% in Vietnam as per published reports.[8],[9],[15],[19] For DHPS, the selection of resistant genotypes is not as restrictive as the DHFR selection.[20] Paired mutations at 436 and 613 appear more frequently in parasite isolates. In a recent study based on in vitro pyrimethamine sensitivity, a predictive hypothesis has been given for characterizing P. falciparum isolates with various DHFR mutations and their antifolate resistance.[21] Parasites exhibiting in vitro Pyr IC50 of above 1000 nM may carry mutations of Asn-108; Ile-51 and / or Arg-59 denote resistance to SP. Like DHFR, there is a correlation between the frequency and number of DHPS mutations with the history and level of Fansidar usage in different geographical areas, which shows evidence to select DHPS-mutant strains after SP treatment. The mutations in DHPS over and above those of DHFR would be predicted to cause SP failure.[8],[9],[22] The sensitivity of the PCR assay in rapid detection of resistant strains makes it an attractive diagnostic method. It is not known to what extent the high correlation between molecular techniques and in vitro drug assay are relevant to the clinical efficacy of SP, but there is an indication of the declining trend of Pyr efficacy. The present study provides information in support of the general hypothesis of associations between various alleles and resistance to drugs in vitro. A systematic controlled clinical trial study on the therapeutic efficacy of SP and antifolate resistance at the epidemiological level may strengthen the utility of this assay. The molecular methods and their application in surveillance programmes to detect drug-resistant mutants may offer an additional tool in epidemiological surveys. Acknowledgements The author is grateful to Dr. Sarala K. Subbarao, Director, Malaria Research Centre, for her encouragement and moral support. The author wishes to thank Dr. Altaf Lal, Chief, Molecular Vaccine Section, CDC, Atlanta, for the kind gift of molecular reagents and Dr. Y. D. Sharma, Professor and Head, Biotechnology, AIIMS, New Delhi for the necessary facilities and support for this study. References
Copyright 2004 - Journal of Postgraduate Medicine The following images related to this document are available:Photo images[jp04006t1.jpg] [jp04006t2.jpg] [jp04006t3.jpg] [jp04006t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}