 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
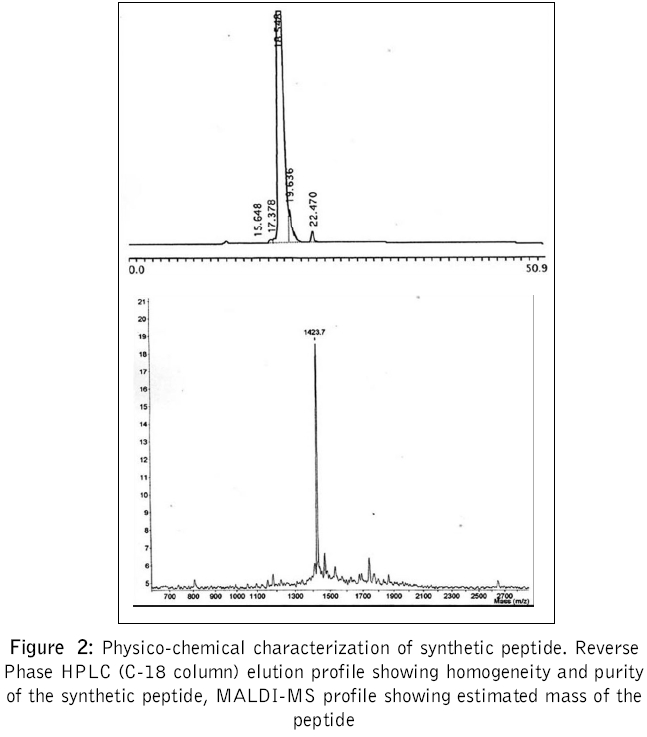
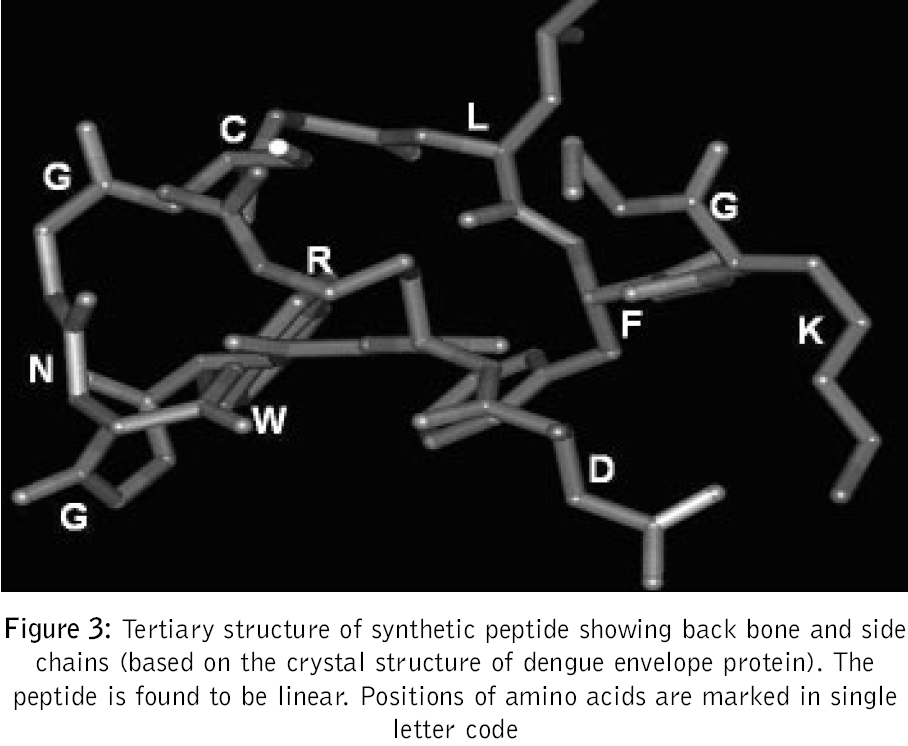
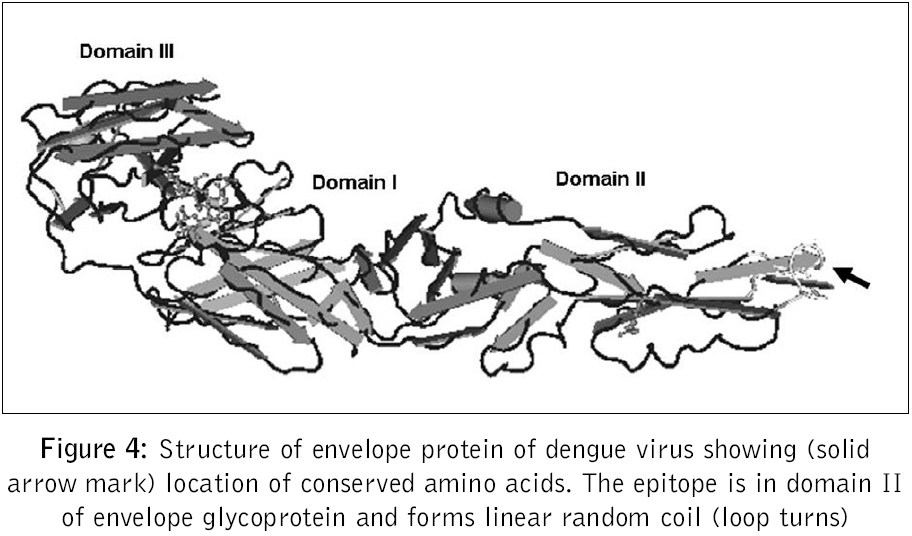
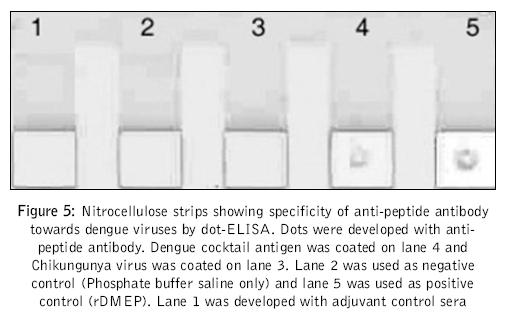
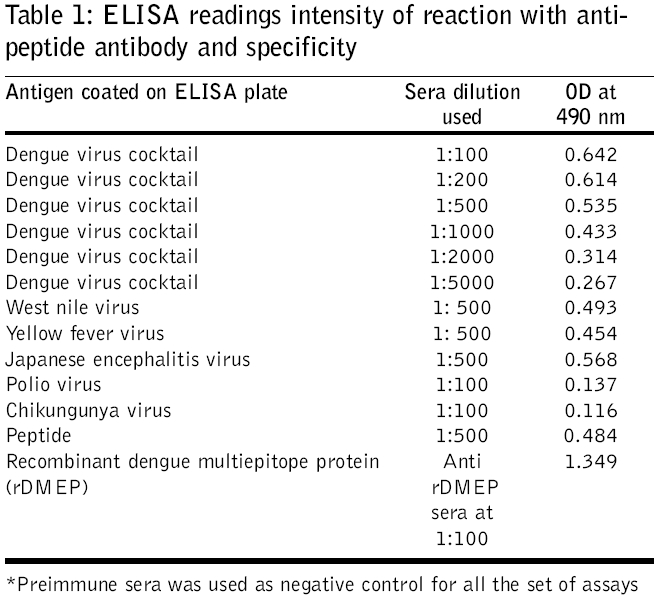
Journal of Postgraduate Medicine, Vol. 52, No. 3, July-September, 2006, pp. 174-178 Original Article Fusogenic peptide as diagnostic marker for detection of flaviviruses Pattnaik Priyabrata, Srivastava A, Abhyankar A, Dash PK, Parida MM, Lakshmana Rao PV Virology Division, Defence Research and Development Establishment, Jhansi Road, Gwalior - 474 002, MP Code Number: jp06059 Abstract Background: Dengue, Japanese encephalitis, West Nile encephalitis, yellow fever are the common flaviviral diseases associated with high morbidity and mortality. The initial symptoms of most of the flaviviral infections are similar to each other as well as to some other viral diseases. Making clinical diagnosis, therefore, becomes a challenging task for the clinician. Several studies have been reported on using detection of serum antibodies against flavivirus for the diagnosis of specific flaviviral disease; no field-based pan-flavi virus detection system is available, which can be used in low-endemicity areas for differentiation of flaviviral disease from other viral diseases.Aim: To identify a conserved amino acid sequence among all flaviviruses and evaluate the antibody formed against the conserved peptide to develop pan-flavivirus detection system. Materials and Methods: In the present study we have compared amino acid sequences of several flaviviruses and identified a conserved amino acid sequence lying in domain II of envelope protein. Results : A peptide having the conserved amino acid sequence was used to generate polyclonal antibodies and these antibodies were used to detect several flaviviruses. Anti-peptide polyclonal antibodies selectively recognized flaviviruses and did not detect non-flaviviruses. Anti-peptide antibodies detected presence of virus in serum spiked with pure virus preparations. Conclusion: The study offers a rationale for development of pan-flavivirus capture assay suitable for low endemic areas. Keywords: Flavivirus, detection, fusion peptide, dot-ELISA, virus capture assay, pan-flavi The last decade witnessed emergence and re-emergence of several microbial infections. A majority of them have been vector-borne infections[1] especially, the arthropod-borne viral diseases, which constitute 90% of the re-emerging diseases. The early manifestations of most of diseases caused by the flaviviruses are very similar to those caused by other viral fevers. From the epidemiological as well as therapeutic point of view, it is essential to differentiate flaviviral infection from other viral infections using a validated laboratory diagnostic method. Several diagnostic systems are available for the detection of circulating antibodies in patient serum.[2],[3] However, no gold standard is available for immunological diagnosis of flaviviruses. Though PCR based diagnosis has become very common, it cannot be used in field conditions and requires trained manpower. A field based simple method to use pan-flavi virus detection system will be of great help specifically for low endemic areas to differentiate flavivirus infections from other viral infections. In the present report we describe an approach for early diagnosis of flaviviral diseases by detecting presence of the viruses using antibody against a peptide corresponding to conserved fusogenic motif of flaviviruses. Materials and Methods Identification of flavi-specific epitope Amino acid sequences of envelope protein of several flaviviruses were downloaded from NCBI databank. Amino acid sequences of Dengue virus type 1 (NCBI accession no. AY153755), Dengue virus type 2 (NCBI accsesion no. M29095), Dengue virus type 3 (NCBI accsesion no. AY099342), Dengue virus type 4 (NCBI accsesion no. AF326573), Japanese encephalitis virus (NCBI accsesion no. NP775666), yellow fever virus (NCBI accession no. NP740305), West Nile virus (NCBI accession no. AAR17573), tick borne encephalitis virus (NCBI accession no. AAC62100), Kyasanur Forest disease virus (NCBI accession no. CAA52211) and Omsk hemorrhagic fever virus (NCBI accession no. NC005062) were considered for evaluation of flavi-specific epitope and identifying conserved amino acid sequences. Amino acid sequences were edited and analyzed by the Lasergene software package V5.0 (DNASTAR Inc, USA). Multiple sequence alignments were carried out employing CLUSTALW version 1.83.[4] Synthesis of peptide The flavi-specific conserved amino acid sequence identified by sequence alignment was custom synthesized from GenoMechanix, L.L.C, Florida, USA. The synthesized peptide was dissolved in triple distilled sterile water so as to achieve a final concentration of 1 mg ml -1 and stored at -20°C till used. Structural and physico-chemical characterization of the peptide The synthesized peptide was analyzed by reverse phase HPLC. Briefly, the synthesized peptide was loaded on a reverse phase C-18 column (Vydac) at a flow rate of 1.5 ml min -1. The gradient used for elution was developed using Buffer A [0.1% trifluoroacetic acid (TFA) in water] and Buffer B (0.1% TFA in 100% acetonitrile). The column was initially equilibrated with Buffer A and reached a final composition of 20% Buffer A and 80% Buffer B in 20 min. MALDI-MS analysis of peptide was done following protocol of Brown and Lennon.[5],[6] Predictive analysis of the peptide Predictive structural analysis of the peptide was carried out using RasWin Molecular Graphics, version 2.7.2.1.1 and Cn3D version 4.1. Structural coordinates of dengue envelope protein (PDB accession no. 1OAN) and Tick borne encephalitis virus envelope protein (PDB accession no. 1SVB) were used for comparative evaluation of peptide for epitope specificity. Antigenicity, hydrophilicity and secondary structure of the peptide was predicted using ProteanTM version 5.07 of Lasergene software package V5.0 (DNASTAR Inc, USA). Animals and immunization Mice used in this study were obtained from the animal facility of the Defence Research and Development Establishment, Gwalior, India. Animals were housed, fed and used in experiments according to the guidelines set forth in the National Institutes of Health manual titled, ′Guide for the care and use of laboratory animals′ (National Institutes of Health Publication No. 86-23, US Department of Health and Human Services, Washington D.C, USA). Swiss Albino mice of 3-4 weeks old were immunized by the subcutaneous route with 100 μg of peptide emulsified in with Freund′s complete adjuvant. The mice were boosted on days 28, 53 and 150 with 100 μg of peptide formulated in Freund′s incomplete adjuvant. Group of five mice were immunized with Freund′s adjuvant alone by the route and schedule described above to provide control anti-serum. Sera were collected 14 days after each immunization and used for immunoassays. ELISA Mice hyper-immune sera were used for recognition of peptide by ELISA. Briefly, wells of a microtitre plate were coated with 1 μg of peptide and blocked with 5% skimmed milk powder solution in phosphate-buffered saline (blocking buffer). Antigen-coated wells were incubated for 90 min at 37°C with 100 μl of mice serum diluted in blocking buffer. Serial two fold dilutions of sera starting with 1:10 were used. After washing with PBS containing 0.05% Tween 20 (Sigma), 100 μl of horseradish peroxidase-labeled anti-mouse IgG antibody conjugate (Sigma) diluted 1:2000 was added to each well and incubated for 60 min at 37°C. The color reaction was developed with o-phenylenediamine dihydrochloride and hydrogen peroxide as the chromogenic substrate. The reaction was stopped by addition of 2M sulfuric acid and OD was recorded at 490 nm in a microplate ELISA reader (BioTek, USA). Pre-immune sera as well as sera raised against adjuvant alone were used with similar dilutions as controls. Dot-ELISA Mice hyper-immune sera were also used for recognition of flaviviruses by dot-ELISA. Briefly, Nitrocellulose strips (NC strips) were coated with 2 μl of heat inactivated virus suspension (~2 μg of total viral protein) of Dengue virus cocktail (all four dengue virus serotypes 1, 2, 3 and 4 in equal proportion), West Nile virus, yellow fever virus, Japanese encephalitis virus respectively. Coated strips were blocked with 2% bovine serum albumin in phosphate-buffered saline (blocking buffer). Antigen-coated strips were kept standing in wells of microtitre plates wells filled with 200 μl of mice serum diluted to 1: 500 in 1% bovine serum albumin in phosphate buffered saline and incubated for 60 min at 37°C. Rest of the procedures are the same as described in ELISA except that the IgG antibody conjugate was used in 7500 dilutions. Mice sera were also tested for recognition of chikungunya virus and polio virus by dot-ELISA in the similar way. Pre-immune sera as well as sera raised against adjuvant alone were used with similar dilutions as controls. NC strips thus developed were dried and observed for appearance of brown color dots. Detection of flavivirus in spiked serum Healthy serum samples separately spiked with purified, heat inactivated dengue virus type-2, West Nile virus, Japanese encephalitis virus and chikungunya virus at concentrations of 10 2, 10 3, 10 4 PFU ml -1 were used to evaluate the specificity of anti-peptide antibody. Wells of a microtitre plate were coated with anti-peptide antibody (serum collected from mice) and blocked with 5% skimmed milk powder solution in phosphate-buffered saline (blocking buffer). Anti-peptide antibody-coated wells were incubated for 90 min at 37°C with 100 ml of spiked healthy serum sample diluted in blocking buffer. After washing with PBS containing 0.05% Tween 20 (Sigma), 100 μl of horseradish peroxidase-labeled IgG purified from dengue patient serum (gratis sample from K. Morita, Nagasaki University, Japan) diluted 1:2000 was added to each well and incubated for 60 min at 37°C. The color reaction was developed with o-phenylenediamine dihydrochloride and hydrogen peroxide as the chromogenic substrate. The reaction was stopped by addition of 2M sulfuric acid and OD was recorded at 490 nm in a microplate ELISA reader (BioTek, USA). A pre-immune serum was used to coat microtire plates as negative controls. OD value twice the OD value of negative control was considered positive. Results Analysis of amino acid sequences of envelope protein of several flaviviruses Clustal W alignment showed a conserved amino acid sequence spanning aa99 to aa112 of envelope protein [Figure - 1]. Conserved amino acid sequence was found to be "DRGWGNGCGLFGKG". The identified peptide was custom synthesized. The purity and homogeneity of the synthesized peptide was analyzed by RP-HPLC, which can separate peptides of even one amino acid difference based on differences in surface hydrophobicity. The synthesized peptide eluted in a single, symmetric peak by reverse phase chromatography on a C-18 column [Figure - 2]A. MALDI-MS analysis was used to assess the mass of the peptide. Mass spectrum of the peptide shows characteristic peak representing a mass of 1423.70 dalton [Figure - 2]B. We performed predictive structural analysis of the peptide to confirm its physical characteristics. The tertiary structure of the peptide in the virus envelope protein was deduced from the reported crystal structure of dengue virus.[7] The peptide backbone with side chains have been shown in [Figure - 3]. The epitope, found to be localized in domain II (A domain) of envelope glycoprotein [Figure - 4], is linear and forms random coil (loop turns). Further predictive analysis using ProteanTM suggested that the peptide is amphiphilic with more hydrophobic residues and may be moderately antigenic. The peptide is found to be easily water soluble and stable upon storage at -20°C. Anti-sera raised in mice with synthetic peptide showed very low titre (1:1500) in ELISA. We performed dot-ELISA using nitrocellulose strips coated with 2 μg peptide per spot. After necessary processing we could see the clear dots when developed with 1:2000 anti-peptide sera dilutions (data not shown). To detect the ability of anti-peptide sera to detect flaviviruses, we did ELISA and dot-ELISA using dengue virus cocktail antigen. In dot ELISA clear dots are visible on NC strips coated with dengue virus cocktail antigen, West Nile virus, yellow fever virus and Japanese encephalitis virus, developed using 1:1000 dilution of anti-peptide sera [Figure 5]. The strips coated with chikungunya virus and polio virus, developed using 1:100 dilution of anti-peptide sera did not yield any visible spots. In plate ELISA, we observed an OD of 0.267 in wells developed with 1:5000 dilutions of anti-peptide sera [Table - 1]. No color development was seen on wells coated with chikungunya virus and polio virus. The anti-peptide sera also selectively recognized 10 3 PFU ml -1 dengue type-2 virus, West Nile virus and Japanese encephalitis virus spiked in healthy serum and did not recognize chikungunya virus spiked at the same concentration in healthy serum. Discussion Dengue fever is one of the arboviral flavivirus infections that is transmitted by mosquitoes, with high morbidity and mortality rate and is extensively prevalent in several parts of tropical countries.[8],[9] The initial symptoms of many flaviviral diseases are overlapping. Though several diagnostic systems are available for detection of presence of antibody in patient serum[2],[3] very little effort has been put to detect flaviviruses during initial viremic phase of infection. Earlier we have reported development of diagnostic test system for detection of circulating antibody in patient serum.[10] Recently our laboratory reported development of a custom-designed recombinant multi-epitope protein for detection of dengue specific IgG from dengue patient serum.[11] In the present study we have used a structural proteomics approach to identify a conserved epitope in envelope protein of several flaviruses and explored possibility to develop flavivirus detection system based on their conserved amino acid sequence. In the present study we used a peptide whose amino acid sequences are conserved among several flaviviruses. The conserved amino acid sequence spans aa99 to aa112 of envelope protein. Except tick borne encephalitis virus (TBEV) and Kyasanur Forest disease virus (KFDV) which has histidine (H), all other viruses have glycine (G) at amino acid position 105. Amino acid position at 98 to 110 has been hypothesized and authenticated in dengue virus that it aids in low pH mediated membrane fusion of virus to mammalian cell membrane.[12] The conserved region we found by homology alignment of amino acid sequences almost overlaps the region regarded as fusion peptide. The custom synthesized peptide elutes in a single, symmetric peak by reverse phase chromatography on a C-18 column suggesting that the peptide is homogenous. Using MALDI-MS, mass of peptide was found to be 1423.70 dalton which is very close to the calculated mass of the peptide, 1422.58 dalton. This analysis confirmed that the synthesized peptide is homogeneous and of desired mass. Predictive structural analysis of the peptide suggested it to be linear, amphiphilic in nature with more hydrophobic residues with moderate antigenicity. This was later confirmed by our results which showed that the peptide is poorly immunogenic and generated low titre antibodies. Anti-sera raised in mice with synthetic peptide showed very low titre (1:1500) in ELISA. Such a low titre of antibody generated against peptide is not uncommon. Several peptides are reported to be non-immunogenic or poorly immunogenic because of either very small size or strong hydrophobicity. The synthesized peptide we used contains more than 50% of amino acids that are hydrophobic and is only of approximately 1.4 kDa in size. This could be the possible reason for its poor immunogenicity. In dot ELISA clear dots are visible on NC strips coated with dengue virus cocktail antigen, West Nile virus, yellow fever virus and Japanese encephalitis virus, developed using 1:1000 dilution of anti-peptide sera [Figure 5]. The strips coated with chikungunya virus and polio virus, developed using 1:100 dilution of anti-peptide sera did not yield any visible spots. These results confirmed that the polyclonal antibody raised against synthetic peptide selectively recognizes dengue virus only and does not recognize chikungunya virus or polio virus, which are non-flavi viruses. The anti-peptide sera also selectively recognized 10 3 PFU ml -1 dengue type-2 virus, West Nile virus and Japanese encephalitis virus spiked in healthy serum and did not recognize chikungunya virus spiked at the same concentration in healthy serum. This observation proves that the anti-peptide antibody has the ability to detect presence of virus in serum samples, hence can be used to develop virus capture assay. Our study indicated that antibody raised against the synthetic peptide corresponding to fusion peptide can be used for detection of flaviviruses. The anti-peptide antibody is also able to detect presence of viruses in spiked serum samples, indicating that it can detect presence of virus in patient serum during early viremic phase of infection. However, it is to be confirmed with patient sera samples showing up at different stages of clinical illness and different stages of viremia. The peptide generates poor antibody response which can be enhanced by coupling to a carrier molecule. Further, monoclonal antibody developed against this synthetic epitope can be used for development of virus capture assay systems/ diagnostic kits that will be of great help for early diagnosis and differential detection of flaviviruses even in the field conditions, specifically for low endemic areas. Acknowledgements We thank Mr. K. Sekhar, Director for his keen interest and support for this work. The technical assistance rendered by Ms. Bharti Agrawal is duly acknowledged. We acknowledge Dr. Subodh Kumar and the reviewer for their critical suggestion on this manuscript.References
Copyright 2006 - Journal of Postgraduate Medicine The following images related to this document are available:Photo images[jp06059f5.jpg] [jp06059f4.jpg] [jp06059t1.jpg] [jp06059f2.jpg] [jp06059f1.jpg] [jp06059f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}