 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
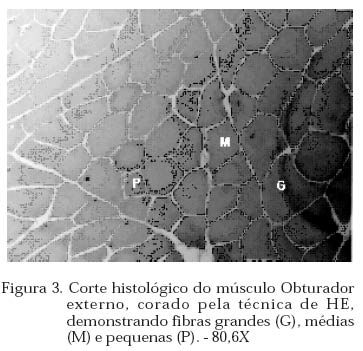
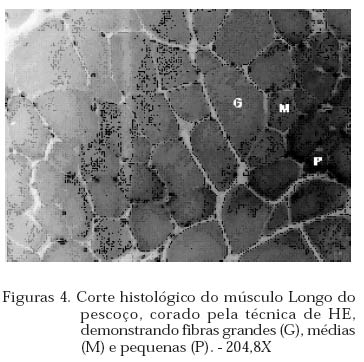
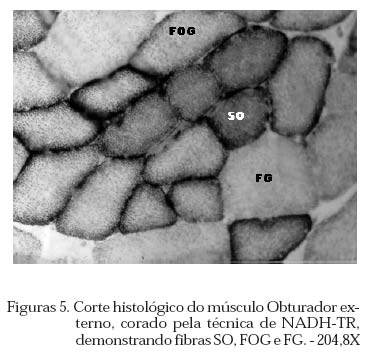
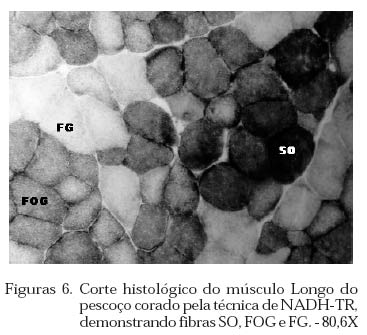
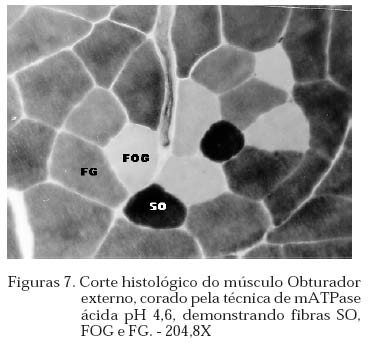
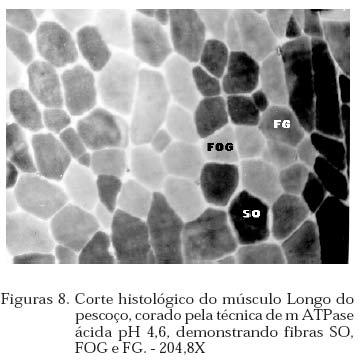
Archivos Latinoamericanos de Produccion Animal, Vol. 12, No. 1, 2004, pp. 1-7 Caracterização dos tipos de fibras de músculos esqueléticos de suínos Landrace, Large White, Duroc e mestiços. Study of the skeketal muscle fibers in purebred and crossbred Landrace, Large White and Duroc swine. S. M. Dierckx 1,J. Bortolozzi 2, V. Dal Pai3 1 Professora Asssitente Doutor - Departamento de Perodução e Exploração Recibido Enero 13, 2002. Code Number: la04001 ABSTRACT Fibers of the external obturador (1) and long of the neck (2) were analyzed in swine, of four genetic group. Histoenzymological techniques were employed (HE, acid mAtpase, pH 4,6 and NADH-TR). This allowed classification as to metabolism type and muscular contraction type. It was concluied that muscle 1 possesses more than 50% of FG fibers and muscle 2 more than 40% of SO fibers. Both presented a larger percentage of fast contraction fibers. Regarding metabolism, muscle 1 has a glycolitic (anaerobic) while 2 has an oxidative (aerobic) character. Key words: NADH-TR, acid mAtpase, HE, fibers So, FOG and FG, swine RESUMO Foram analisadas fibras dos músculos Obturador esterno (1) e Longo de pescoço de suínos, de quatro grupos genéticos. As técnicas histoenzimológicas (HE, mAtpase ácida, pH 4,6 e NADH-TR) permitiram classificá-los quanto ao tipo de metabolismo (anaeróbico e aeróbico) e de contração muscular (lenta e rápida). Os resultados permitiram concluir que o músculos 1 possui mais de 50% de fibras FG e o 2 mais de 40% de fibras SO e que ambos apresentaram maior percentagem de fibras de contração rápida e quanto ao metabolismo o 1 tem caráter glicolítico e o 2 oxidativo. Palavras chave: NADH-TR, mAtpase ácida, HE, fibras SO,FOG,FG, suínos. Introdução A parte comestível dos suínos é principalmente, tecido muscular estriado esquelético que constitui os músculos. A estrutura básica do músculo é a fibra muscular; um conjunto delas forma um feixe muscular e um conjunto de feixes forma o músculo, envolvido pelo tecido conjuntivo. Roça e Serrano (1994) afirmam, que as funções vitais do sistema muscular não cessam no momento da morte do animal; ao contrário, as modificações bioquímicas e estruturais, que ocorrem após o sacrifício, são chamadas de «conversão do músculo em carne». Elas ocorrem simultaneamente e são dependentes dos tratamentos ante-mortem, do processo de abate e das técnicas de armazenamento da carne. No decorrer das três ultimas décadas, o domínio progressivo das condições de criação, associado à seleção dos animais, baseada no aumento da velocidade de crescimento, na redução do estado de adiposidade e na melhora da eficiência alimentar, permitiram ampliar, consideravelmente, a produtividade do rebanho suíno. Porém, o impacto, ao longo prazo, sobre as características musculares dos suínos, foi por muito tempo desprezado. Foram principalmente os dois tipos de carnes «problema» PSE («pale, soft, exudative») e DFD (dark, firm dry or dark cutting) que evidenciaram a necessidade de levar em conta, não somente a quantidade de músculos, mas também suas características qualitativas. Peter et al. (1972), propôs a classificação das fibras musculares em três tipos, colocados abaixo, levando em conta a velocidade de contração e o tipo de metabolismo: SO: contração lenta, metabolismo aeróbico, baixa atividade glicolítica e de moderada a alta atividade aeróbica. FOG: contração rápida, metabolismo oxidoglicolítico, moderada a alta atividade anaeróbica, alto teor de citocromos e atividade oxidativa. FG: contração rápida, metabolismo glicolítico, alto teor de glicogênio, baixo teor de citocromos, e baixa atividade oxidativa. A distinção entre fibras FOG e FG pode ser realizada somente a partir de quatro semanas pós-parto (Lefaucheur et al., 1983). Depois a conversão FOG - FG se opera principalmente na periferia das ilhas de fibras Slow e traduz a passagem de um metabolismo óxido-glicolítico para um metabolismo glicolítico. O tamanho de um músculo, isto é a quantidade de carne, depende do número de fibras musculares, de seu tamanho (comprimento, diâmetro) e da quantia de tecido conjuntivo. No suíno, o número total de fibras (NTF) é fixado desde antes do nascimento (Stickland e Goldspink, 1973). O potencial de crescimento muscular devido ao NTF depende, então, do período pré-natal, porque o crescimento pós-natal se faz por adição de núcleos novos (hiperplasia), dentro das fibras a partir das células satélites indiferenciadas, por ampliação celular (hipertrofia) Close, (1972), e por acúmulo de tecido conjuntivo. O número de fibras musculares é fixo e característico nas diferentes raças e linhagens genéticas de suínos (Staun, 1963; Davies, 1972). Dubowitz (1968), Romanul (1964), Close (1972), afirmam que os músculos vermelhos, ou seja, aqueles que possuem maior porcentagem de fibras vermelhas, apresentam metabolismo aeróbico, contração lenta e atividade prolongada e com predominância de fibras brancas têm atividade de contração descontínua e de curta duração. É de grande importância conhecer a microestrutura do tecido muscular para se traçarem planos nutricionais e de melhoramento visando a produção de carne em termos de qualidade e quantidade. Ferrari (1994) comentando os resultados obtidos na citação Handel e Stickland (1988) em animais Large White, estudados com o objetivo de estabelecer uma possível relação entre o potencial de crescimento e o número de fibras musculares esqueléticas em animais que nascem com alto ou baixo peso corporal, positivou que o número de fibras musculares ao nascimento representa um indicador do crescimento pós-natal dos suínos. Staun (1963) pesquisando o músculo Longissimus dorsi de suínos das raças Landrace de linhagem alemã, Pietrain, Duroc, Large White e outras duas raças locais alemãs, concluiu que nos animais, machos inteiros, castrados e fêmeas, de 100kg não houve diferenças significativas para o número e o diâmetro das fibras. Staun (1972) afirma que, nos trabalhos de seleção para aumento da carne em suínos, o emprego das mensurações entre indivíduos pode ser eficiente, mas, o estudo de amostras de músculos será de grande valia para encontrar uma melhor qualidade da carne produzida. As modificações na estrutura muscular levam a alterações na qualidade da carne produzida, uma vez que a seleção visando animais com alto peso muscular, tem levado igualmente a se selecionar, músculos com maior número de fibras musculares por grupos metabólicos e a aumentar a quantidade de fibras musculares de contração lenta O presente trabalho teve como objetivo a análise histoenzimológica dos músculos Obturatori externi e Longus colli, visando:- verificar se haveriam diferenças entre as raças estudadas, quanto ao tipo de fibras musculares, caracterizar os músculos quanto ao tipo de contração muscular e caracterizar os músculos quanto ao tipo de metabolismo. Materiais e Métodos O estudo foi realizado com 12 suínos com peso ao redor de 90 kg, oriundos da região de Botucatu, sendo 3 Landrace, 3 Large White, 3 Duroc e 3 mestiços, de ambos os sexos abatidos em frigorífico. As amostras foram coletadas na linha de abate do frigorífico, no momento em que as carcaças eram serradas ao meio, sendo o manejo antes e pós abate o rotineiramente adotado. Foram utilizadas amostras de 0,5cm de comprimento por 0,5cm de largura, dos músculos Obturatori externi (Obturador externo) e Longus colli (longo do pescoço) (Figuras 1 e 2), passadas em talco e colocadas em nitrogênio líquido, até o momento das análises, como usual para as análises histoenzimológicas. Processamento do material Os fragmentos musculares foram transferidos para câmara de criostato Keichert Jung CM 1800, à -20oC, repousando por uma hora, de acordo com Pullen (1977). A seguir, as amostras musculares foram fixadas aos suportes metálicos com adesivo O.C.T. (Optimal Critical Temperature), e os blocos foram orientados de maneira a se obterem cortes transversais ao maior eixo das fibras musculares. Para cada músculo de cada um dos animais, foram feitas três séries de cortes com 10mm de espessura. A primeira série foi usada para a coloração por hematoxilina e eosina (HE) (Lillie, 1954) para a descrição dos aspectos morfológicos dos músculos. A segunda série foi usada para a demonstração da atividade da Nicotinamida Adenina Dinucleotídeo Tetrazólium Reductase (NADH-TR), de acordo com o método de Pearse (1968), modificado por Dubowitz e Brooke (1973), permitindo a classificação e o cálculo das porcentagens dos diferentes tipos de fibras musculares. A última série foi submetida à reação para o estudo da atividade enzimática da Adenosina Trifosfato miofibrilar (mATPase) ácida em pH 4,6, à temperatura ambiente por 15 minutos e pré-incubação em pH 10,2, segundo Padykula e Herman (1955). As fibras musculares foram classificadas de acordo com a nomenclatura de Peter et al. (1972) identificando os três tipos de fibras: SO, FOG e FG. Classificação, contagem e percentagem das fibras musculares A classificação dos diferentes tipos de fibras musculares foi feita nos cortes histológicos submetidos às reações NADH-TR e mATPase, após pré-incubação em pH ácido. Utilizou-se «uma câmara clara», acoplada a um microscópio ZEISS, visualizando 10 (dez) campos microscópicos, de cada animal, com objetiva 40X e ocular 10X. Análise estatística O delineamento experimental adotado foi o inteiramente casualizado, incluindo no modelo estatístico os efeitos de Grupo Genético, Animal/Grupo Genético, Músculo e Músculo/Grupo Genético. As porcentagens de fibras SO, FOG e FG foram convertidas em arco seno √x e a análise de variância foi efetuada no Sas (1985). As médias foram comparadas pelo teste de Tukey, segundo Gomes (1973). Os cortes histológicos, corados com hematoxilina e eosina (HE), permitiram verificar que ambos os músculos se apresentavam divididos em feixes musculares envolvidos por tecido conjuntivo, com os núcleos localizados geralmente na porção periférica das fibras musculares (Figuras 3 e 4). Pela análise dos cortes histológicos usados para o estudo das reações enzimáticas para mATPase e NADH-TR, pré incubada em pH ácido (4,6), (Figuras 5 , 6, 7, a 8), pode-se notar que ambos os músculos, apresentavam os três tipos de fibras musculares (SO, FOG e FG), e as fibras musculares encontravam-se distribuídas em «grupos metabólicos», com as SO mais centrais e distribuídas ao seu redor as fibras FOG. 3.1 Tipo das fibras musculares SO, FOG e FG 3.1.1. Total das fibras e número de fibras SO, FOG e FG O número total de fibras musculares não foi significativamente influenciado por grupo genético, animal/grupo genético músculo e músculo/grupo genético. Discorda dos achados de Davies (1972) e Stickland & Goldspink (1973) e concorda com Staun (1963). O total de fibras SO e FG foi influenciado significativamente apenas pelos tipos de músculos (P<0,01). O músculo Obturador externo apresentou 48,83 fibras musculares SO a menos que o longo do pescoço e 88,51 fibras FG a mais.(Quadro 1) 3.1.2. Percentagens das fibras SO, FOG e FG Os músculos apresentaram diferenças na percentagem das fibras PSO (P<0,01), PFOG (P<0,05) e PFG (P<0,01). As médias obtidas para as percentagens de fibras SO, FOG e FG (Quadro 2) evidenciam que, os músculos não diferiram entre si, quanto à percentagem de fibras FOG, mas sim, quanto à percentagem de fibras SO e FG. O músculo 1 apresentou menor percentagem de fibras FG e o 2 de fibras SO. Os resultados obtidos quando comparados aos de Ferrari (1994), que estudando o músculo Longissimus dorsi de suínos Duroc obteve 13,60% de fibras SO, 17,25% de FOG e 69,67% de fibras FG, evidenciam que os músculos aqui, estudados têm comportamento diferente. O músculo 1 possui 21,84%, 24,35% e 53,81% de fibras SO, FOG e FG e o 2, respectivamente: 43,47%, 28,97% e 27,57%. Quadro 1. Médias estimadas do número de fibras SO e FG, em relação aos músculos Obturador externo (1) e Longo do pescoço (2)
Médias seguidas de letras distintas na coluna diferem estatisticamente entre si a 5% pelo teste de Tukey 3.2. Comparação quanto ao tipo de metabolismo oxidativo (SO + FOG) e glicolítico (FG) 3.2.1. Número de fibras (SO+FOG) e FG Para o estudo do tipo de metabolismo foram agrupados as fibras SO + FOG e FG, os resultados obtidos permitiram concluir, que o efeito de músculo foi a única causa de variação que influenciou significativamente (P<0,01) o tipo de metabolismo oxidativo e glicolítico. As médias do número de fibras (SO+FOG) e FG nos dois músculos foram comparadas e estão no Quadro 3. Ela exibe que os músculos diferem entre si apenas quanto ao número de fibras de metabolismo glicolítico (FG). Assim o músculo 1 apresentou 88,51 fibras FG a mais que o 2, que apresentou 50 fibras de metabolismo oxidativo a mais que o 1. Nota-se também que os músculos não diferiram entre si quanto ao metabolismo oxidativo. O músculo 1 não diferiu quanto ao número de fibras de metabolismo oxidativo e glicolítico; ao contrário do músculo 2, que exibiu 124,85 fibras de metabolismo oxidativo a mais. 3.2.2. Percentagem das fibras de metabolismo oxidativo (PSO+PFOG) e glicolítico (PFG) Procedeu-se também ao estudo do tipo de metabolismo oxidativo ou glicólico, agrupando-se as percentagens de fibras SO e FOG, obtendo-se que apenas os músculos influenciaram a percentagem de fibras musculares de metabolismo oxidativo (PSO+PFOG) e glicolítico (PFG). Isso sugere que os músculos têm diferentes tipos de metabolismo. As médias obtidas para as percentagens de fibras musculares de metabolismo oxidativo e glicolítico, nos dois músculos foram comparados pelo teste de Tukey e encontram-se no Quadro4. Observa-se pelo Quadro 4 que o músculo 1 possui maior percentagem de fibras do metabolismo glicolítico que do tipo oxidativo, embora a diferença obtida entre os dois grupos seja pequena, enquanto o 2 apresenta 72,433% de fibras de metabolismo oxidativo e apenas 27,567% de fibras de metabolismo glicolítico. 3.3. Estudo do tipo de contração muscular 3.3.1. Análise do tipo de contração muscular baseada no número de fibras musculares de contração rápida (FOG+FG) e lenta (SO) Com a finalidade de se verificar o tipo de contração rápida ou lenta, efetuaram-se as análises de variância que mostraram que somente os músculos influenciaram (P<0,01) o número de fibras musculares de contração rápida (FOG+FG) e lenta (SO), indicando que, os músculos apresentaram diferentes tipos de contração. As médias obtidas para o número de fibras musculares de contração rápida (FOG+FG) e lenta (SO), nos dois músculos foram comparadas pelo teste de Tukey e encontraram-se no Quadro 5. Esta indica que o músculo, 1 possui 87,34 fibras musculares de contração rápida a mais que o 2, e que ao contrário, possui 48,83 fibras musculares de contração lenta a mais, em relação ao outro. Quadro 2. Médias estimadas das percentagens de fibras SO (PSO), FOG (PFOG) e FG (PFG), nos músculos Obturador externo (1) e Longo do pescoço (2)
Médias seguidas de letras distintas na coluna diferem estatisticamente entre si a 5% pelo teste de Tukey Quadro 3. Médias estimadas do tipo de metabolismo oxidativo (SO +FOG) e glicolítico (FG), nos músculos Obturador externo (1) e Longo do pescoço (2)
Médias seguidas de letras distintas na coluna diferem estatisticamente entre si a 5% pelo teste de Tukey Quadro 4. Médias estimadas das percentagens de fibras de metabolismo oxidativo (PSO+PFOG) e glicolítico (PFG), nos músculos Obturador externo (1) e Longo do pescoço (2)
Médias seguidas de letras maiúsculas distintas na coluna diferem estatisticamente entre si a 5% pelo teste de Tukey Os músculos possuem maior número de fibras de contração rápida (P<0,05), embora a diferença seja muito maior no número 1. 3.3.2. Análise do tipo de contração muscular rápida e lenta baseada nas percentagens de fibras (PFOG+PFG) e PSO Com a finalidade de conhecer qual o tipo de contração utilizando-se as percentagens de fibras de contração rápida (FOG+FG) e lenta (SO), foram efetuadas as análises de variâncias que permitiram notar que apenas os músculos influenciaram (P<0,05) a percentagem de fibras musculares de contração rápida e lenta, e que novamente indica, possuírem diferentes tipos de contração muscular. As médias obtidas para as percentagens de fibras musculares de contração rápida (PFOG+PFG) e len ta (PSO), nos músculos estudados, foram comparadas pelo teste de Tukey e fixam-se no Quadro 6. O Quadro 6 indica que ambos os músculos possuem contração rápida (P<0,05). O músculo 1 apresenta uma alta percentagem (78,160) de fibras de contração rápida. No número 2 a diferença entre a percentagem de fibras musculares de contração rápida é bem menor (13,07%), indicando que o músculo 2 possui alta porcentagem de fibras musculares de contração lenta. Os estudos histoenzimológicos dos músculos estudados possibilitaram verificar, que eles apresentaram diferenças quando ao aspecto externo (tamanho, forma, localização, cor) e tiveram também diferentes percentagens de fibras SO, FOG e FG. O músculo 1 apresentou mais de 50% de fibras FG e o 2 mais de 40% de fibras SO. Para a comparação do tipo de contração muscular, observou-se, que ambos os músculos apresentaram maior percentagem de fibras de contração rápida, apesar desta «característica» ser mais evidente no músculo 1. Para a comparação do tipo de metabolismo, obteve-se que o músculo 1 possuía maior percentagem de fibras de metabolismo glicolítico; mas, no músculo 2 a diferença entre a percentagem de fibras de metabolismo oxidativo foi bem maior que a das fibras de metabolismo glicolítico. Quadro 5. Médias estimadas do número de fibras de contração rápida (FOG+FG) e lenta (SO), nos músculos Obturador externo (1) e Longo do pescoço (2)
Médias seguidas de letras distintas na coluna diferem estatisticamente entre si a 5% pelo teste de Tukey Quadro 6. Médias estimadas das percentagens de fibras de contração rápida (PFOG+PFG) e lenta (PSO), nos músculos Obturador externo (1) e Longo do pescoço (2)
Médias seguidas de letras maiúsculas distintas na coluna diferem estatisticamente entre si a 5% pelo teste de Tukey Quando se analisam os resultados histoenzimológicos, nota-se que, os músculos apesar de possuírem diferentes percentagens de fibras SO e FG, apresentaram maior percentagem de fibras de contração rápida, diferindo apenas quanto ao tipo de metabolismo. O músculo 1 apresentou mais fibras do tipo glicolítico e o 2 do tipo oxidativo. Os resultados também detectaram não haver diferenças para os grupos genéticos. 1. Não houve diferenças entre os grupos genéticos. 2. As técnicas histoenzimológicas, mostraram haver diferença entre os músculos quanto às percentagens de fibras SO e FG 3. Os dois músculos apresentaram maior percentagem de fibras de contração rápida embora a diferença seja mais acentuada para o músculo1 4. Quanto ao tipo de metabolismo o músculo 1 apresentou maior percentagem de fibras de metabolismo glicolítico e o 2 de metabolismo oxidativo. Literatura Citada
© 2004 ALPA. Arch. Latinoam. Prod. Anim. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}