 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 12, No. 1, 2004, pp. 12-20 Efeito da sustituição do farelho de soja pela levedura (Saccharomyces cerevisiae) desidratada como fonte protéica em dietas para leitões desmamados sobre peso de órgãos digestivos e atividade das enzimas pancreáticas. Effect of the substitution of soybean meal by dehydrated yeast (Saccharomyces cerevisiae) as protein source in diets to weaned piglets on the weight of digestive tract organs and activity of pancreatic Enzymes W. Castillo1, R. N. Kronka, J. M. Pizauro Jr., M. C. Thomaz, L. E. Carvalho2. 1Bolsista da Fundação de Amparo à Pesquisa
do Estado de São Paulo, FAPESP. Dirección actual: Facultad de
Zootecnia, Universidad Nacional Agraria de la Selva,Tingo María, Perú.
E-mail: wilicaso@hotmail.com Recibido Septiembre 2, 2002. Code Number: la04003 ABSTRACT The effect of protein-equivalent substitution of soybean meal by spray-dry dehydrated yeast (SDDY) on the weight and development of digestive organs and activities of pancreatic enzymes was evaluated. Sixty 21-day old, individually housed, weaned piglets were distributed according to a randomized blocks design with a 5 x 4 factorial arrangement (5 dietary substitution levels: 0, 25, 50, 75, and 100% x 4 time periods: 0, 4, 8, and 12 d after weaning). SDDY substitution level did not influence daily bodyweight gain and weight of stomach; a quadratic response was found in daily dry matter intake (DMI), as described by the equation: Y = 133.3 + 1.466X 0.0178X2 (R2 = 0.99) and in weight of small intestine: Y = 287.2 + 0.697X 0.01115X2 (R2 = 0.99), estimated best results being obtained with 41% of substitution; whereas weight of pancreas decreased linearly (P < 0.01). Trypsin activity was not affected but lipase and amylase activity decreased in a quadratic (P < 0.05) fashion, respectively, with increasing SDDY in the diet. It may be concluded that during these post-weaning periods a decrease in pancreatic protein content and pancreatic growth rate associated with the low DMI found in the first days post weaning, may limit the synthesis of pancreatic enzymes. Key Words: Digestive organs, pancreatic enzymes, early-weaned pigs, yeast. RESUMO Foi avaliado o efeito produzido pela substituição do farelo de soja (FS) pela levedura desidratada em «spray dry» (LDSD) como fonte protéica sobre o desempenho, o desenvolvimento dos órgãos digestivos e a atividade das enzimas digestivas pancreáticas. Foram utilizados 60 leitões desmamados aos 21 dias de idade, alojados individualmente e distribuídos num delineamento de blocos ao acaso com arranjo fatorial 5x4 (5 níveis de substituição dietética: 0, 25, 50, 75 e 100% x 4 épocas de abate: 0, 4, 8 e 12 dias após o desmame). A LDSD não influenciou a ganância diária de peso e o peso do estômago; encontrou-se respostas quadráticas para o consumo diário de ração - CDR (Y = 133.299 + 1.466X - 0.0178X2; R2 = 0.99) e para o peso do intestino delgado (Y = 287.196 + 0.697X - 0.01115X2; R2 = 0.99), conseguindo-se melhores resultados com 41% de substituição; entanto que, o peso do pâncreas decresceu linearmente (P<0.01). A atividade da tripsina não foi afetada, enquanto da lipase e da amilase decresceram de forma quadrática (P<0.01) e linear (P<0.05), respectivamente, com o aumento da LDSD na dieta. Avaliando os períodos pós desmame, pode-se concluir que a diminuição no conteúdo de proteína e na taxa de crescimento do pâncreas, associado ao baixo CDR encontrado nos primeiros dias após o desmame, estariam limitando a síntese das enzimas pancreáticas. Palavras Chave: Leitões, Levedura, Enzimas pancreáticas, Órgãos digestivos Introdução Nos primeiros dias após o desmame dos leitões, os órgãos como pâncreas, estômago e intestino delgado tendem a variar de peso, bem como, a diminuição na atividade enzimática, sendo essa variação dependente da idade ao desmame (Efird et al., 1982b), da suplementação de nutrientes (Kelly et al., 1991a), do tipo de fonte protéica na dieta (Makkink et al., 1994; peiniau et al., 1996) ou da presença direta de nutrientes no lúmem (Remillard et al., 1998). As alterações observadas na primeira semana após o desmame, acompanhadas de uma redução no desempenho e no sistema enzimático pancreático nem sempre são recuperadas por volta da segunda semana. O farelo de soja (FS) usado nas rações, pode apresentar fatores antinutricionais insuficiente inativados, proteínas antigênicas podem se encontrar no íleo dos leitões e não serem hidrolisadas pelas enzimas proteolíticas, devido a que, estas podem ter a sua atividade comprometida (Armour et al., 1998). A avaliação de outras fontes protéicas, como a levedura desidratada (Saccharomyces cerevisiae), obtida no processo de fermentação alcoólica, pela industria brasileira, após termólise e secagem pelo processo «spray dry» (LDSD) com teor médio de 40% de proteína bruta, pode ser alternativa de uso como substituto do FS na alimentação de suínos. Para determinar se a LDSD influencia o desenvolvimento do sistema digestivo e o desempenho de leitões após o desmame foi desenvolvido um experimento, tendo como objetivo avaliar a substituição da proteína do FS pela proteína da LDSD sobre o desenvolvimento dos órgãos digestivos e a atividade das enzimas digestivas pancreáticas (lipase, amilase e tripsina). Material e Métodos O Experimento foi conduzido no setor de Suinocultura, no Laboratório de Nutrição Animal e no Laboratório de Enzimologia Aplicada da Faculdade de Ciências Agrárias e Veterinárias, UNESP, Câmpus de Jaboticabal - SP, Brasil. Foram utilizados 60 leitões híbridos comerciais, de ambos sexos, desmamados aos 21 dias de idade, com peso médio de 5.90 + 0.75 kg. Após o desmame foram abatidos 15 leitões, sendo os outros alojados em baias individuais onde receberam as mesmas condições de manejo, bem como dieta farelada e água à vontade até os dias de abate. Os tratamentos foram o resultado da combinação dos fatores, níveis de substituição da PB do FS pela PB da LDSD na dieta (0, 25, 50, 75 e 100%) e épocas de abate dos leitões (0, 4, 8 e 12 dias após o desmame). As dietas foram isoprotéicas e isocalóricas (Tabela 1) e formuladas atendendo as exigências nutricionais dos leitões na fase inicial, seguindo as normas do NRC (1988). Foi avaliado o desempenho (ganho diário de peso - GDP e consumo diário de ração - CDR) em cada período de abate; desenvolvimento do estômago, pâncreas e intestino delgado (peso total e peso relativo - g/kg de peso vivo); atividade das enzimas digestivas (expresso em unidades de atividade enzimática (UA), lipase e amilase (UA/mg de proteína e UA/g de pâncreas), tripsina (UA/g de proteína e UA/g de pâncreas) e, conteúdo de proteína pancreática (mg/g de pâncreas). Amostragem e determinação da Atividade das Enzimas Digestivas e da Proteína Os leitões foram deixados em jejum de 15 a 16 h. Ao abate, eles foram insensibilizados e praticada uma incisão na linha alba, sendo o pâncreas retirado e pesado, estômago e intestino delgado foram lavados e pesados. Os pâncreas foram congelados em nitrogênio líquido e armazenados a - 70 ºC. Todo este processo foi realizado em aproximadamente 15 min. Para a extração dos zimogênios e das enzimas ativas, cada pâncreas congelado foi triturado em um moínho. Alíquotas foram homogeneizadas em um homogeneizador OMNI GLH utilizando-se solução salina (NaCl 0.9%) contendo triton X-100 0.1%. O homogeneizado foi centrifugado a 14000 g por 30 min a 4 ºC, e o sobrenadante obtido foi filtrado em lã de vidro e recuperado. O extrato bruto do pâncreas (EBP) assim obtido foi utilizado na determinação da atividade das enzimas digestivas. Alíquotas do EBP foram congelados em nitrogênio líquido e armazenadas a -70 ºC para dosagem de proteína. Para determinar a atividade da lipase (EC 3.1.1.3) foi necessário obter a colipase parcialmente purificada a partir de extrato de pâncreas delipidado, seguindo o procedimento descrito por Brockman (1981). A atividade da lipase foi determinada, para cada amostra em triplicata, por titulometria segundo o procedimento descrito por Sarda e Desnuelle (1958). Usou-se como substrato emulsão de óleo de oliva (SIGMA 800 -1) contendo TRIS.HCl 0.2 M, pH 8.0, taurodeoxicolato de sódio 6 mM, NaCl 0.15 M, CaCl2.2H2O 1 mM e colipase de suíno parcialmente purificada. O meio foi mantido sob agitação a 37 ºC e interrompida a reação com etanol 95%. Os ácidos graxos liberados foram determinados por titulação com NaOH. A atividade da tripsina (EC 3.4.21.4) foi determinada logo da ativação do tripsinogênio presente no EBP através da incubação por 30 min do extrato bruto com enteroquinase em tampão TRIS.HCl 0.5 M, pH 8.0, contendo CaCl2.2H2O 50 mM. A atividade da tripsina determinou-se em triplicata, seguindo o procedimento descrito por Kakade et al. (1974), tendo como substrato o N-a-benzoil-DL-arginina-p-nitroanilida (BAPNA), contendo TRIS.HCl 50 mM, pH 8.0 e CaCl2.2H2O 0.2 M. A reação foi incubada a 37 ºC e interrompida com ácido acético 30%, centrifugado (aproximadamente 100 000 g) por 1 min sob refrigeração e a p-nitroanilida liberada foi determinada a 410 nm (e = 9620 M-1 cm-1) em um espectrofotômetro HITACHI U-2000. Controles foram incubados para determinar a presença de tripsina na solução de enteroquinase bem como a hidrólise espontânea do substrato. Tabela 1. Composição percentual e valores calculados das dietas para leitões na fase inicial. Substituição do FS pela LDSD (%)1
1 Substituição feita a nível
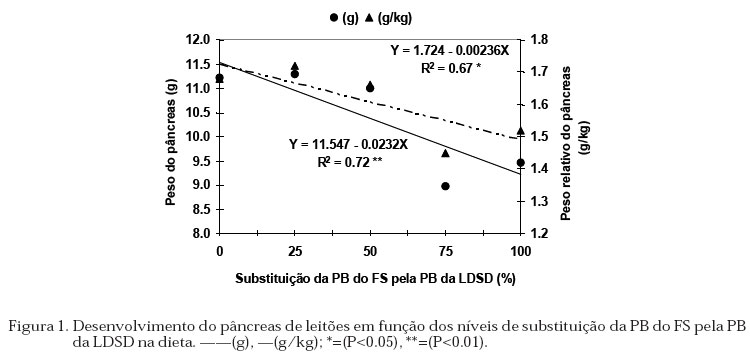
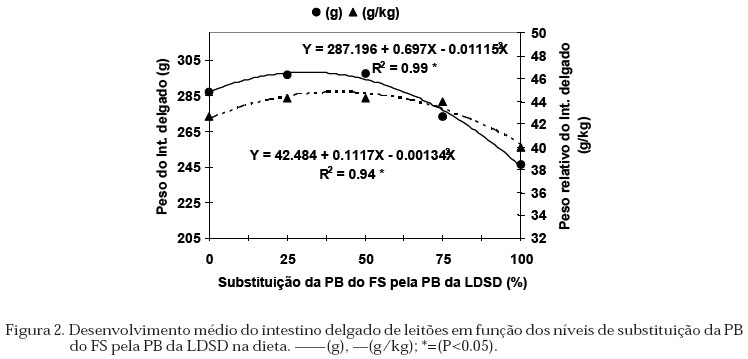
protéico. A atividade da a-amilase (EC 3.2.1.1) pancreática foi determinada com cinco replicações, através da dosagem da maltose liberada a partir da hidrólise do substrato (amido) pela enzima presente no EBP e nas condições padrões do teste (tampão fosfato 25 mM, pH 6.9, NaCl 3.3 mM e amido solúvel 1%; Oser, 1965). Após incubação a 37 ºC a reação foi interrompida com o reagente do ácido dinitrosalicílico contendo, ácido 3,5-dinitrosalicílico 0.04 M, NaOH 0.4 N e tartarato duplo de sódio e potássio 1.06 M. As amostras foram colocadas em banho-maria em ebulição, resfriadas, diluídas com água destilada, homogeneizadas e a absorbância determinada a 530 nm. Foram realizadas dosagens com concentrações conhecidas de maltose, para determinar uma reta padrão e, também foram incluídos controles para se estimar a hidrólise não enzimática do substrato. As Unidades de Atividade enzimática (UA) foram calculadas, para lipase: 1 mmol de ácido graxo liberado/min; tripsina: 1 mmol de p-nitroanilida liberada/min e; amilase: 1 mmol de maltose liberado/min. O conteúdo de proteína do extrato de pâncreas foi determinado com cinco replicações, pelo método descrito por Hartree (1972), usando-se soroalbumina bovina como padrão. Delineamento Experimental e Análise Estatística Foi utilizado o delineamento em blocos casualizados, em esquema fatorial 5 x 4 (5 dietas x 4 épocas de abate), com 3 blocos, sendo o peso dos animais ao desmame o fator de bloqueio e tendo um animal como parcela. Os dados foram submetidos à análise de variância e análise de regressão polinomial, utilizando o programa ESTAT (Sistema de Análise Estatística - UNESP/FCAVJ). Resultados e Discussão Desempenho dos Leitões e Desenvolvimento dos Órgãos Não foi encontrada interação significativa entre os níveis de substituição da PB do FS pela PB da LDSD e os períodos avaliados para GDP e CDR. O efeito dos níveis de inclusão da levedura são mostrados na Tabela 2, observou-se que estes não influenciaram o GDP, porém o CDR apresentou uma resposta quadrática (Y = 133.299 + 1.466X - 0.0178X2; R2 = 0.99), mostrando o maior consumo com o valor calculado de 41.18% de substituição da PB do FS pela PB da LDSD. A não diferencia significativa no GDP entre os animais que receberan diferentes níveis de levedura foi influenciada pela grande variação no GDP obtida nos leitões nos primeiros cuatro dias após o desmame, neste período alguns leitões até perderam peso, resultando em um mayor SEM. O maior CDR de animais que consumiram dietas contendo até 13% de levedura (aproximadamente 52% de substituição) foi atribuído à melhora na palatabilidade (Landell filho, 1991 e Miyada et al., 1992). Porém, a variação encontrada no CDR influenciado pelos níveis de levedura não afetaram o GDP dos animais, concordando com os resultados observados por Makkink et al. (1994) os quais encontraram aos 3 dias após o desmame maior CDR nos leitões que receberam dieta contendo farelo de soja e farinha de peixe em relação àqueles que consumiram leite desnatado em pó, atribuindo à forma física da dieta contendo leite desnatado em pó como a responsável do menor CDR. No desenvolvimento dos órgãos, não observou-se interação significativa entre os níveis de levedura e os períodos avaliados para os pesos dos órgãos. A inclusão da LDSD na dieta não influenciou o peso do estômago (Tabela 2), enquanto os pesos do pâncreas (total e relativo) decresceram linearmente (Figura 1) e os pesos do intestino tiveram resposta quadrática em função dos níveis de levedura na dieta (Figura 2). Quanto ao peso do estômago, tem-se atribuído ao CDR e à quantidade de energia consumida como sendo os fatores diretos que influenciam a variação. Porém, os resultados obtidos não mostraram essa correspondência com o CDR, discordando com os observados por Kelly et al. (1991b). Também tem sido demostrado que as fontes protéicas não afetam o peso do estômago (Makkink et al., 1994) e que animais que consumiram leite após o desmame em quantidade para suprir só a energia de mantença apresentaram estômagos menos pesados em relação àqueles que consumiram ração ou leite à vontade (Pluske et al., 1996). Isto sugere que para o peso do estômago, mais importante que o tipo de proteína que é fornecido, é a atenção das necessidades nutricionais dos animais. Tabela 2. Médias de ganho de peso, consumo de ração e peso do estômago (g e g/kg de peso vivo) dos leitões, segundo os níveis de substituição da levedura, avaliados nos períodos após o desmame.
1 Níveis de substituição da proteína do FS pela proteína da LDSD
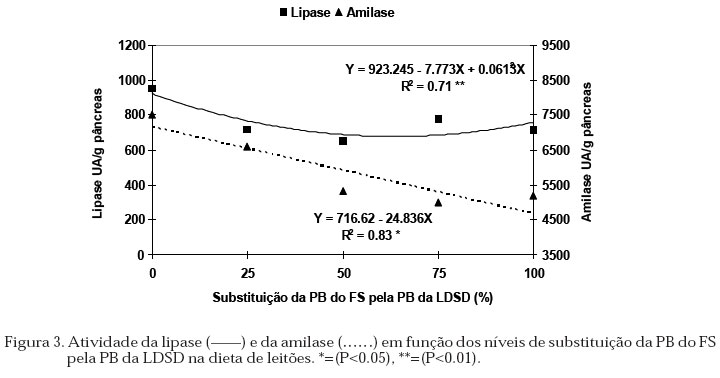
O peso do pâncreas em relação ao peso vivo, foi afetado pelo nível de levedura. A proporção na diminuição do peso do pâncreas com o aumento dos níveis de levedura na dieta não foi a mesma do peso relativo (Figura 1), sugerindo que os animais tiveram pâncreas mais leves em relação ao peso corporal quando comparado àqueles que não receberam levedura, consequentemente pesos relativos maiores indicaram pâncreas mais pesados em relação ao peso corporal. Os resultados obtidos no peso do pâncreas foram semelhantes aos obtidos por Efird et al. (1982a) e Lindemann et al. (1986) os quais observaram que a adição de produtos lácteos às dietas induziu hipertrofia do pâncreas. Pâncreas com pesos relativos maiores foram observados em leitões desmamados aos 14 dias de idade, alimentados durante 7 dias, com dieta contendo leite desnatado em pó, farinha de peixe e farelo de soja quando comparado àqueles que receberam leite materno (Kelly et al., 1991a). A adição de farelo de soja ou farinha de peixe às rações de leitões, ocasionou aumento no peso do pâncreas respectivamente aos 3 e aos 10 dias após o desmame, quando comparado com aqueles que receberam concentrado protéico de soja ou leite desnatado em pó (Makkink et al., 1994). Peiniau et al. (1996) relataram que as propriedades físico-químicas da proteína como a solubilidade associada à digestibilidade como sendo as responsáveis pelo desenvolvimento do pâncreas. Animais que receberam concentrado protéico solúvel de peixe tiveram menor peso do pâncreas em relação àqueles que receberam caseína, farelo de soja ou concentrado protéico de soja dos 21 aos 56 dias de idade. Estes fatos sugerem uma resposta direta do desenvolvimento do pâncreas em função do consumo de alimento, das fontes protéicas e das características físicas das mesmas. No presente experimento, a solubilidade não foi avaliada, devendo relacionar o desenvolvimento do pâncreas com os níveis de levedura nos tratamentos experimentais. No intestino delgado (Figura 2), as variações nos pesos podem estar associadas com o consumo de alimento. As respostas aos níveis de levedura sobre o CDR e o peso do intestino foram quadráticas, obtendo-se o maior CDR, maior peso total e peso relativo do intestino com os níveis calculados de 41.18, 31.25 e 41.67% de levedura respectivamente. As relações de CDR e peso relativo do intestino após o desmame foram relatados por Kelly et al. (1991b), Makkink et al. (1994) e Zijlstra et al. (1996). Pluske et al. (1996) determinaram que a quantidade de energia que os leitões consomem após o desmame é a responsável pela variação no peso do intestino, demonstrando que existe um componente de adaptação no desenvolvimento do intestino associado com a quantidade de alimento consumido. Quando foram avaliados os períodos (do desmame até 12 dias após), os órgãos mostraram uma resposta linear de aumento no peso com a idade (Tabela 3). O aumento do peso do estômago (total e relativo) estão de acordo com as determinações de Lindemann et al. (1986), Kelly et al. (1991a) e Makkink et al. (1994) com leitões desmamados aos 28, 14 e 28 dias, respectivamente. Para o intestino delgado, verificaram diminuição do peso relativo nos 3 dias seguintes ao desmame, e atribuíram o efeito ao menor consumo de alimento nesse período quando comparado ao consumido antes do desmame. Neste experimento, embora os leitões tenham apresentado aos 4 dias seguintes ao desmame, semelhante CDR que os reportados por Makkink et al. (1994), os pesos do pâncreas e do intestino delgado aumentaram linearmente com a idade. Atividade das Enzimas Pancreáticas Os níveis de substituição da PB do FS pela PB da LDSD e os períodos avaliados não mostraram interação significativa para nenhuma das enzimas estudadas nem para o conteúdo de proteína do extrato de pâncreas. Avaliando o efeito da levedura pela análise de regressão sobre as enzimas encontrou-se que, tanto o conteúdo de proteína no pâncreas como a atividade da tripsina não foram influenciados pela levedura na dieta (Tabela 4). A atividade da lipase, quando expressa em UA/g pâncreas mostrou uma resposta quadrática, enquanto que a atividade da amilase decresceu linearmente com o aumento da levedura na dieta (Figura 3). Os valores de atividade, quando expressos em UA/mg de pâncreas, foram semelhantes aos obtidos por Lindemann et al. (1986) para as enzimas lipase e amilase e por Owsley et al. (1986) para a tripsina, determinados em leitões de 21 e 27 dias, respectivamente, indicando adequada padronização das metodologias neste experimento. A síntese e secreção das enzimas pancreáticas tem mostrado respostas diferentes às fontes protéicas. Assim, tem sido verificado que o uso do soro de leite (Lindemann et al., 1986), leite desnatado em pó e o concentrado protéico de soja (Makkink et al., 1994) aumentaram a atividade da tripsina nos primeiros dias após o desmame. Todavia, o concentrado protéico solúvel de peixe apresentou menor atividade total da tripsina e amilase em relação à caseína, farelo de soja e concentrado de soja (Peiniau et al., 1996). A influência imediata e de maior importância fisiológica exercida pelos ingredientes das rações parece ser sobre a secreção das enzimas pancreáticas no lúmem intestinal. Efird et al. (1982a) e Owsley et al. (1986) encontraram diferentes níveis de atividade da tripsina no conteúdo intestinal de suínos alimentados com dietas contendo leite ou soja, entretanto, a atividade no pâncreas e a atividade total permaneceram constantes. As afirmações anteriores foram confirmadas por Valette et al. (1992) que encontraram influência direta na secreção pancreática pela fonte protéica da dieta em leitões canulados no duto pancreático. A correlação entre consumo diário de ração e atividade enzimática foi relatado por Makkink et al. (1994), salientando o efeito da fonte protéica da dieta sobre o estimulo da síntese e secreção enzimática. Neste experimento, o maior CDR (estimado em 41.18% de substituição da PB do FS pela PB da LDSD) ocasionou menor atividade da lipase, valores intermediários da amilase (Figura 3) e sem afetar na atividade da tripsina (Tabela 4), discordando dos dados apresentados por Makkink et al. (1994). Tabela 3. Médias de peso dos órgãos digestivos (g e g/kg de peso vivo) nos leitões, segundo os períodos avaliados após o desmame.
1 Efeitos da inclusão da levedura pela
análise de
Regressão:. L Efeito Linear, * = (P<0.05), ** = (P<0.01). Tabela 4. Atividade enzimática (UA) da tripsina e conteúdo de proteína no tecido pancreático dos leitões, segundo os níveis de substituição da levedura, avaliados nos períodos após o desmame.
1 Níveis de substituição
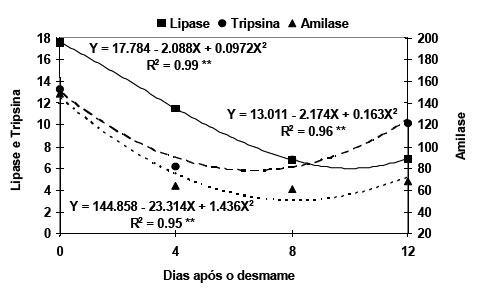
da proteína do FS pela proteína da LDSD Avaliando o efeito dos períodos estudados, para o conteúdo de proteína do extrato de pâncreas verificou-se efeito cúbico, quando expresso em mg/g de pâncreas (Y = 159,253 - 32,026X + 4,554X2 - 0,198X3, R2= 1.0, P<0.01), diminuindo com o tempo após o desmame. Para as atividades da lipase, tripsina e amilase os efeitos foram quadráticos, independente da forma de expressão (Figura 4). As enzimas apresentaram comportamento diferente quanto ao número de dias para atingir a mínima atividade. A lipase e amilase começaram a se recuperar depois dos 10.7 e 8.11 dias, respectivamente. A tripsina apresentou queda mais rápida, tendo atividade mínima aos 6.7 dias, sendo que aos 12 dias mostrou valores semelhantes aos obtidos no dia do desmame. Os resultados obtidos estão de acordo com os reportados por Efird et al. (1982b), Lindemann et al. (1986) e Owsley et al. (1986) os quais determinaram os valores de atividade das enzimas em função da idade do desmame. A queda na atividade da lipase pode ser atribuída ao fato de que os lipídios do alimento, que os leitões ingerem, diminuem de cerca de 40% da matéria seca do leite materno para menos de 5% na dieta na fase após o desmame. No caso da amilase, os altos valores de atividade encontrados no desmame foram semelhantes aos determinados em leitões desmamados com 4 semanas e que receberam durante a lactação dieta para leitões (Shields et al., 1980) ou ingeriram a dieta da mãe (Lindemann et al., 1986). A tripsina mostrou alta atividade no dia do desmame. A queda e a rápida recuperação ocorreu em função da mudança do tipo de proteína que os leitões ingeriram. Efird et al. (1982b), Owsley et al. (1986), Lindemann et al. (1986) e Makkink et al. (1994) enfatizaram que a quimotripsina é a enzima proteolítica predominante na lactação, enquanto que a tripsina incrementa rapidamente após o desmame com a mudança do alimento. A diminuição no conteúdo de proteína e na taxa de crescimento do pâncreas, associado ao baixo CDR que caracteriza os primeiros dias após o desmame, estariam limitando a síntese das enzimas por reduzir os níveis de precursores disponíveis (Owsley et al., 1986) e por comprometer o estímulo que exercem os nutrientes a nível molecular para a expressão gênica das enzimas (Corring et al., 1989). Conclusão A substituição da proteína do farelo de soja pela proteína da levedura até 41% melhorou o consumo da dieta e o peso relativo do intestino, sem afetar o ganho diário e o peso do estômago. Porém, uma substituição total, influenciou negativamente o peso do pâncreas, provocou diminuição na atividade da lipase e amilase e não afetou a atividade da tripsina. Na avaliação pós desmame, se verificou que a diminuição no conteúdo de proteína e na taxa de crescimento do pâncreas, associado ao baixo CDR encontrado nos primeiros dias após o desmame, estariam limitando a síntese das enzimas pancreáticas. Literatura Citada
© 2004 ALPA. Arch. Latinoam. Prod. Anim. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}