 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 12, No. 3, Septiembre-Deciembre, 2004, pp. 96-102
Parâmetros séricos de vacas leiteiras na fase inicial de lactação
Serous metabolite levels of early lactation dairy cows
S. E. López, J. López, W. Stumpf Junior
Departamento de Zootecnia da Faculdade de Agronomia da Universidade Federal do Rio Grande do Sul, Puerto Alegro RGS. Brasil
Recibido Enero 30, 2003 Code Number: la04012
Abstract. Eight Jersey cows in early lactation with mean liveweight of 380 kg were distributed into two 4 x 4 latin squares with 21-day experimental periods, and fed corn silage, alfalfa hay, and four concentrates (control and with three different sources of fat: tallow, commercial protected fat, and ground whole soybeans, so as to provide 6.3% EE in the total dietary DM vs. 3.7% in the control). The objective was to evaluate effects of the lipid supplementation on blood composition. Blood samples were collected on days 17 and 21 of each experimental period via jugular vein puncture using vacutainer tubes with a separator gel, followed by centrifugation. The sources of fat did not influence serous levels of glucose, the enzyme GGT, and Ca; however, differences (P<0.05) were found in the serous concentrations of cholesterol, triglycerides, non-esterified fatty acids and urea. The values found in this study are within normal ranges according to the literature on serous levels of the evaluated metabolites. Key Words: Blood Metabolites, Dairy Cows, Protected Fat, Soybean, Tallow. Resumo. Foram utilizadas oito vacas da raça Jersey, no início da lactação, com peso vivo médio de 380 kg distribuídas em dois quadrados latinos 4 x 4 com período experimental de 21 dias, e alimentadas com silagem de milho, feno de alfafa e quatro rações concentradas (controle e três contendo diferentes fontes de gordura: sebo, gordura protegida comercial e grãos de soja integral triturados de modo a alcançar 6.3 % de EE na MS da dieta total vs. 3.7% no controle). O objetivo foi avaliar o efeito da suplementação lipídica sobre os parâmetros sangüíneos. As coletas de amostras de sangue foram realizadas aos 17 e 21 dias de cada período experimental através de punção da veia jugular e recolhidas em tubos vacutainer com gel separador, sendo após centrifugadas. As fontes de lipídeos não afetaram (P>0.05) os níveis séricos de glicose, enzima g-glutamil-transferase e cálcio, enquanto houve diferença (P<0.05) nas concentrações séricas de colesterol, triglicerídeos, ácidos graxos não esterificados e uréia entre os tratamentos. Os níveis séricos dos metabólitos avaliados encontram-se dentro da faixa de normalidade quando comparados aos valores citados na literatura consultada.
Palavras-chave: Gordura Protegida, Grão de Soja, Metabólitos Sangüíneos, Sebo, Vacas Leiteiras.
INTRODUÇÃO
Em anos mais recentes nutricionistas têm procurado formular dietas que supram as exigências energéticas de vacas leiteiras no início da lactação, período este em que os animais, especialmente de alta produção, apresentam-se em balanço energético negativo, pois não conseguem consumir a quantidade necessária de alimento para suprir suas exigências nutricionais. Os lipídeos têm sido usados como forma de aumentar a densidade energética da dieta (em média contêm 6.5 Mcal/kg de EB), sem contudo alterar a relação volumoso : concentrado ou aumentar o incremento calórico. Além de minimizar o balanço energético negativo as gorduras previnem desordens metabólicas e melhoram o desempenho na lactação e na reprodução, além de restaurar a condição corporal. Várias fontes de lipídeos podem ser utilizadas como o sebo animal, as sementes de oleaginosas tais como o algodão, o girassol e a soja e a gordura protegida. Esta última fonte de gordura apresenta a vantagem adicional de não influenciar negativamente a fermentação ruminal, pois é submetida a técnicas industriais de proteção. O conhecimento das variações na concentração sangüínea de metabólitos como glicose, colesterol, triglicerídeos, ácidos graxos não esterificados, enzima g-glutamil-transferase, uréia e cálcio ajuda a explicar as respostas zootécnicas e metabólicas observadas mediante o fornecimento de lipídeos na dieta de vacas em lactação (Gagliostro e Chilliard, 1992). O objetivo deste estudo foi avaliar os efeitos da utilização de dietas sem e com suplementação de diferentes fontes de gordura sobre os parâmetros séricos de vacas Jersey no início da lactação.
MATERIAL E MÉTODOS
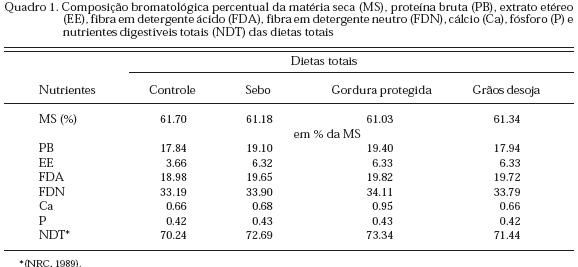
Foram utilizadas oito vacas puras por origem da raça Jersey, pertencentes ao rebanho leiteiro da EMBRAPA-CPACT localizada em Capão do Leão - RS, multíparas, com peso corporal médio de 380 kg, com escore corporal de 3.5 a 4.0 segundo classificação de Wildman et al. (1982) e diferentes dias de parto, mas aproximados. Os animais se encontravam na 2a ou 3a ordem de lactação com uma média de 38 dias em lactação. Antes de iniciar o experimento, todos os animais foram pesados e depois colocados em baias individuais. O teste de OPG (contagem de ovos de nematóides por grama de fezes) apresentou resultado negativo. Os tratamentos testados foram os seguintes: Tratamento 1 - Concentrado sem a inclusão de fonte de gordura; Tratamento 2 - Concentrado + sebo bovino; Tratamento 3 - Concentrado + gordura protegida e Tratamento 4 - Concentrado + grãos de soja integral triturados. Foi utilizado no experimento um galpão tipo «free stall», onde as baias estavam divididas por meio de cordas e sendo utilizados sarrafos para mantê-las esticadas. A higienização das baias e dos bebedouros foi feita diariamente. Os concentrados foram compostos de grãos de milho moídos, farelo de soja, fosfato bicálcico, pré-mistura mineral e vitamínica e calcário, cujo fator de variação foi a inclusão ou não de diferentes fontes de gordura. Estas gorduras foram adicionadas em certos níveis de modo a tornar as dietas suplementadas isoenergéticas, não deixando de levar em consideração as exigências estabelecidas para vacas leiteiras pelo NRC (1989). A gordura protegida de nome comercial «Enerjet» é composta por 100 % de óleos vegetais constituídos por ácidos graxos de óleo de palma de alta digestibilidade protegidos em um pH 6.2 por ser um sabão cálcico, conforme indicado no rótulo pelo fabricante. A composição química, segundo as informações contidas no mesmo rótulo é a seguinte: gordura bruta = 84 %; cinzas = 12-13 %; umidade = 3-4 % e energia metabolizável = 6.5 Mcal. Os volumosos utilizados no experimento foram o feno de alfafa obtido de um estabelecimento comercial e a silagem de milho produzida em silos-trincheira próximos ao «free stall». A formulação das dietas foi realizada com auxílio de um programa de computador e a dieta foi calculada com o objetivo de manter a proporção de volumoso e concentrado de 60:40, respectivamente. No Quadro 1 é observada a composição bromatológica das dietas totais. O concentrado foi fornecido individualizado por animal no cocho de alimentação primeiramente e em separado do volumoso, após cada ordenha e também, uma vez, no intervalo de tempo entre as duas ordenhas do dia. Foram misturados 30 g de bicarbonato de sódio no concentrado antes de ser colocado no cocho. Não foram observadas sobras do concentrado. Os volumosos foram fornecidos para cada vaca duas vezes ao dia (após cada ordenha) em quantidades previamente pesadas, mas de modo a que sempre houvesse sobras para garantir o seu consumo à vontade. Foram feitas amostras compostas de concentrados e volumosos para cada tratamento. Água fresca e de boa qualidade ficou permanentemente à disposição dos animais em tonéis plásticos. As vacas foram ordenhadas mecanicamente, duas vezes ao dia, às 7h e 30 min e às 17h e 30 min. As análises das amostras de volumosos e concentrados foram realizadas segundo técnicas descritas na AOAC (1996); os teores de fibra em detergente neutro (FDN) e fibra em detergente ácido (FDA) foram determinados conforme método de Goering e Van Soest (1970). Os teores de cálcio dos alimentos e leite foram determinados por espectrofotometria de absorção atômica conforme método de Tedesco et al. (1995). As concentrações séricas de glicose, colesterol, triglicerídeos, uréia, cálcio e enzima g-glutamil-transferase foram determinadas pelo método enzimático-colorimétrico utilizando kits comerciais. Os ácidos graxos não esterificados do soro sangüíneo foram dosados pelo método enzimático-colorimétrico de Johnson e Peters (1993) também utilizando um kit comercial.
Durante a última semana de cada período experimental, no 17o e 21o dia, foram coletadas, antes da refeição da manhã e após a ordenha, três amostras de sangue de cada animal para as determinações dos componentes séricos. A coleta das amostras de sangue foi feita através de punção na veia jugular, após a contenção do animal. Para a retirada do material foram utilizados tubos vacutainer com gel separador. À medida em que as amostras foram coletadas, os tubos foram colocados em uma estante de tubos dentro de uma caixa isotérmica fechada. Tendo sido encerrada a coleta, as amostras foram imediatamente centrifugadas para a obtenção do soro. Como delineamento no experimento foram utilizados dois quadrados latinos 4 x 4, sendo cada quadrado composto por quatro vacas, quatro dietas e quatro períodos experimentais, cada um com duração de 21 dias (Pimentel Gomes, 1990). Os dados foram analisados utilizando o programa estatístico SAS (SAS, 1998). As variáveis foram analisadas pela análise de variância dos valores médios de cada tratamento e as médias pelo teste F.
RESULTADOS E DISCUSSÃO
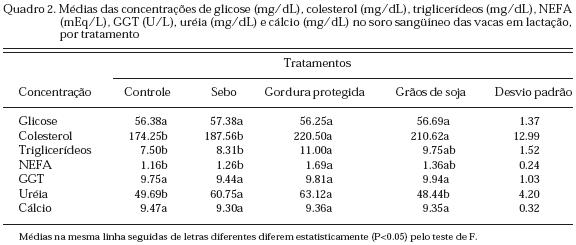
No Quadro 2 podem ser observados os valores médios referentes à concentração de glicose, colesterol, triglicerídeos, ácidos graxos não esterificados (NEFA), enzima hepática g-glutamil-transferase (GGT), uréia e cálcio no soro sangüíneo das vacas em lactação por tratamento. Na análise de variância das concentrações de glicose no soro sangüíneo não foi observada diferença significativa para fonte de gordura utilizada ou não. Não era esperado efeito dos tratamentos sobre a concentração deste metabólito, pois segundo Gagliostro e Chilliard (1992), diversos mecanismos de economia de glicose pelo organismo dos ruminantes podem explicar a manutenção da glicemia apesar da redução no consumo de MS, que ocorre muito freqüentemente ao utilizar-se a suplementação lipídica: diminuição na oxidação da glicose para produzir NADPH necessário para a lipogênese de novo, devido a uma inibição da mesma nos tecidos adiposo e mamário frente ao aporte de lipídeos; diminuição da oxidação da glicose para produção de ATP, que pode ser produzido a partir da oxidação dos ácidos graxos exógenos; possível aumento na gliconeogênese hepática, como conseqüência de uma menor concentração plasmática de insulina e um aumento da concentração do hormônio do crescimento e, finalmente, uma eventual resistência à insulina do organismo suplementado com lipídeos, que poderia contribuir para manter a glicemia. As variações nas concentrações de glicose sérica de bovinos vão de 35.0 a 55.0; 45.0 a 75.0 e 42.1 a 74.5 mg/dL segundo Blood e Radostits (1991); Clinical Biochemistry of Domestic Animals (1997) e Manual Merck de Veterinária (1997), respectivamente. Os resultados encontrados no presente experimento situam-se dentro daqueles campos de variação. O efeito da suplementação lipídica sobre a concentração de glicose no sangue nos trabalhos revisados foi muito variada, pois existem aqueles onde não houve alteração no nível sangüíneo (Gagliostro, 1997; Bermudes, 1999; Abdullah et al., 2000), enquanto outros observaram aumento no teor de glicose sangüínea ao fornecerem suplementação lipídica (Elliott et al., 1993; Bateman et al., 1996). Estes últimos autores, por exemplo, verificaram um aumento, em média, de 57.58 mg/dL para 63.60 mg/dL de glicose no plasma para os grupos controle e suplementados com sebo, respectivamente. A análise de variância das concentrações de colesterol no soro sangüíneo mostrou diferença significativa para a fonte de gordura utilizada ou não, sendo que os animais que receberam a gordura na forma protegida ou como grãos de soja triturados apresentaram maiores (P<0.05) teores deste parâmetro em relação aos demais tratamentos, mas não diferiram entre si (P>0.05). As variações nas concentrações de colesterol no soro de bovinos vão de 39 a 177 e 62.1 a 192.5 mg/dL nas publicações de Blood e Radostits (1991) e Manual Merck de Veterinária (1997), respectivamente. Uma série de estudos têm mostrado respostas hipercolesterolêmicas em vacas em lactação consumindo suplementos lipídicos (West e Hill, 1990; Drackley e Elliott, 1993; Elliott et al., 1993). Por exemplo, West e Hill (1990) obtiveram um aumento no teor de colesterol sérico de 221.74 mg/dL para 300.02 mg/dL, ao adicionar sais cálcicos de ácidos graxos à dieta de vacas leiteiras; do mesmo modo, Drackley e Elliott (1993), comparando dietas com níveis crescentes de sebo parcialmente hidrogenado (0, 2, 4 e 6 % na MS total da dieta), observaram um aumento na concentração de colesterol no plasma (176; 230; 247 e 250 mg/dL) ao suplementarem os animais. Segundo Nestel et al. (1978), o aumento na concentração de colesterol no sangue em vacas consumindo dietas com suplementação de lipídeos deve-se a um aumento na síntese intestinal de colesterol necessário para a absorção e transporte de elevados teores de lipídeos circulantes provindos da dieta e também a uma redução na excreção fecal de ácidos biliares. Mesmo não tendo sido medido o conteúdo de colesterol no leite do presente experimento, é interessante observar que pesquisadores como Wrenn et al. (1976); Wrenn et al. (1977) e Rafalowski e Park (1982), verificaram que apesar do aumento na concentração de colesterol no sangue com a suplementação lipídica, o seu conteúdo no leite não aumentou e também não houve alteração no seu conteúdo na carne (Wrenn et al., 1977). Nos trabalhos de Wrenn et al. (1976) e Wrenn et al. (1977), foi observado que a transferência do colesterol do «pool» sangüíneo para o leite foi limitada. Este fato, segundo os mesmos autores, indica que o colesterol não é transferido diretamente do sangue para o leite, pois existem «pools» metabólicos diferentes para o colesterol do sangue e do leite. Conduzindo uma série de estudos para investigar a origem do colesterol do leite no ruminante, Long et al. (1980) constataram que embora uma certa quantidade de colesterol seja sintetizado através da síntese de novo na glândula mamária, ele é derivado principalmente da sua fração presente no soro sangüíneo. Os dados destes estudos também indicaram que o colesterol sérico equilibra-se com o colesterol das membranas das células mamárias lactantes antes da sua transferência e secreção no leite. Estas observações parecem significar que a concentração de colesterol no leite está mais estreitamente relacionada ao seu conteúdo no «pool» da membrana do que à sua concentração sangüínea (Rafalowski e Park, 1982). A análise de variância das concentrações de triglicerídeos no soro revelou diferença estatística para a adição ou não de uma fonte de gordura à dieta. Os animais dos tratamentos que receberam gordura protegida ou grãos de soja triturados apresentaram maior concentração sérica de triglicerídeos (P<0.05); todavia, este último tratamento não diferiu (P>0.05) dos demais. Segundo os quadros do Clinical Biochemistry of Domestic Animals (1997) os valores séricos normais de triglicerídeos para bovinos situam-se entre 0 e 14 mg/dL e os valores deste trabalho se enquadram nesse campo de variação. A elevação dos níveis de triglicerídeos sangüíneos foi freqüentemente observada em trabalhos onde foram estudados os efeitos da suplementação lipídica à dieta (Smith et al., 1978; Chow et al., 1990) e, provavelmente, reflete o aumento na digestão e captura de ácidos graxos a partir da gordura fornecida (Avila et al., 2000). Chow et al. (1990), por exemplo, obtiveram um aumento de 7.65 mg/dL para 12.65 mg/dL no plasma de vacas em lactação, ao adicionarem gordura amarela à dieta. No entanto, o aumento na concentração deste metabólito é fraco e não sistemático devido à sua alta taxa de troca (Gagliostro e Chilliard, 1992). Contrariando estes autores, West e Hill (1990) e Gagliostro (1997), trabalhando com sais cálcicos de ácidos graxos; Bermudes (1999), utilizando gordura protegida de origem marinha e Kronfeld et al. (1980), testando sebo protegido, não obtiveram diferença ao adicionarem estes lipídeos à dieta. Na análise de variância das concentrações de ácidos graxos não esterificados (NEFA) no soro foi observada diferença estatística para a inclusão ou não de uma fonte de gordura à dieta, sendo que os animais dos tratamentos que receberam suplementação de gordura protegida ou grãos de soja triturados apresentaram maiores níveis deste parâmetro (P<0.05); entretanto, este último tratamento não diferiu dos demais (P>0.05). O escalonamento dos dados encontrados para os NEFA acompanha pari-passu o que se observa com os triglicerídeos no soro sangüíneo. Existe muita controvérsia entre os autores em relação ao efeito da suplementação lipídica sobre os teores de NEFA no sangue de vacas em lactação. A literatura mundial apresenta vários trabalhos onde a adição de gordura provocou um aumento na concentração dos NEFA sangüíneos (Bertics et al., 1999; Avila et al., 2000). Gagliostro (1997), por exemplo, adicionando ou não 400 g de sais cálcicos de ácidos graxos para vacas em lactação, observou um aumento na concentração dos NEFA de 378 para 446 mEq/L com a suplementação. Entretanto, outros autores não verificaram alteração no teor deste metabólito no sangue (Pires et al., 1996; Bermudes, 1999). Segundo Grummer e Carroll (1991), o aumento nos níveis dos NEFA na corrente sangüínea de vacas em lactação suplementadas com lipídeos deve-se a uma incompleta captura destes pelos tecidos após a hidrólise dos triglicerídeos de lipoproteínas de muito baixa densidade (VLDL) pela lipoproteína lipase e/ou a um aumento na hidrólise de triglicerídeos do tecido adiposo. A análise de variância não mostrou diferença estatística significativa para fonte de gordura suplementada ou não na concentração da g-glutamil-transferase (GGT), sendo que a média geral correspondeu a 9.74 U/L. Esta enzima se caracteriza por sua extrema sensibilidade. No caso de alterações hepáticas, o aumento verificado na sua concentração servirá como um índice de agressão tóxica (Kramer e Hoffmann, 1997). Os resultados obtidos no presente trabalho indicaram que os teores de gordura das dietas não comprometeram a atividade hepática e renal dos animais. Bermudes (1999), estudando o efeito da inclusão ou não de gordura protegida na dieta de vacas em lactação, não observou diferença entre os tratamentos em relação ao teor da GGT plasmática dos animais, com uma média geral de 31 UI/L. Na análise de variância das concentrações de uréia no soro foi observada diferença estatística para a suplementação ou não de gordura às dietas. Os animais que consumiram a gordura na forma protegida ou de sebo apresentaram maiores (P<0.05) teores de uréia sérica em relação aos animais dos outros dois tratamentos. Kim et al. (1993) observaram que as concentrações de uréia sérica e amônia ruminal foram menores para as dietas suplementadas com grãos de soja extrusados (17 % na MS da dieta total) ou sais cálcicos de ácidos graxos (4 % na MS da dieta total). Os autores concluíram que esta redução indicou uma utilização mais eficiente dos carboidratos fermentáveis ruminalmente para a síntese de proteína microbiana ou uma inibição na atividade proteolítica microbiana. Outros autores não encontraram diferença entre tratamentos em relação ao teor de uréia sangüínea ao testar diferentes fontes de gordura (Grum et al., 1996; Gagliostro, 1997). Testando a adição ou não de diferentes fontes de gordura à dieta (controle, grãos de soja tostados, sais cálcicos e sebo hidrogenado), Dhiman et al. (1995) não encontraram diferença entre os tratamentos quanto ao teor de uréia no plasma dos animais (51.7; 50.2; 51.8 e 52.7 mg/dL, respectivamente). As concentrações de uréia no sangue e no leite estão altamente correlacionadas. A observação do Quadro 2 e os valores da concentração de uréia obtidos no leite (T1-31.70; T2-35.88; T3-37.72 e T4-31.18 mg/dL) permitem confirmar esta afirmativa. Elas podem ser utilizadas como um indicador da utilização e adequação do fornecimento de nitrogênio para vacas em lactação. Esta pressuposição está baseada na concepção de que os conteúdos de uréia no sangue e no leite podem fornecer informações a respeito das perdas de N após a absorção de amônia a partir do trato gastrointestinal, particularmente o rúmen, segundo Oltner et al. (1985). Estes autores encontraram uma alta correlação (r=0.91) entre a uréia do leite e a do sangue. Existe um grande interesse prático na busca por parâmetros de fácil execução que ajudem a estimar ou avaliar a adequação do fornecimento de nutrientes para vacas leiteiras no sentido de manter uma alta produção de leite e boa saúde. Um possível indicador seria a concentração de uréia no leite, que está altamente correlacionada com a relação proteína : energia da dieta (Oltner e Wiktorsson, 1983) e serve também para investigar problemas de fertilidade (Oltner et al., 1985). Este parâmetro tem a vantagem de evitar o estresse da coleta de sangue, além de que a amostragem é de fácil obtenção e o processo de análise do leite é simples, rápido e de baixo custo (nas condições dos Estados Unidos) (Oltner e Wiktorsson, 1983). A determinação da concentração de uréia no leite não pode substituir a análise convencional do alimento, mas sim, pode servir como um complemento importante. Quanto ao cálcio não seria de esperar-se uma maior variação na sua concentração desde que sua homeostase é quase perfeita face aos vários mecanismos que a controlam. A análise de variância não foi significativa para a inclusão ou não de uma fonte de gordura em relação ao parâmetro cálcio sérico, sendo que a média geral correspondeu a 9.37 mg/dL. As variações nas concentrações de cálcio no soro de bovinos são de 8.0 a 10.5; 9.7 a 12.4 e 8.4 a 11.0 mg/dL nas publicações de Blood e Radostits (1991); Clinical Biochemistry of Domestic Animals (1997) e Manual Merck de Veterinária (1997), respectivamente. Os valores encontrados para o cálcio sérico no presente trabalho (média de 9.37 mg/dL) se situam dentro da variação da concentração do cálcio, segundo estes autores. Kronfeld et al. (1980); Mohamed et al. (1988) e West e Hill (1990), também não encontraram diferença na concentração de cálcio sangüíneo ao testarem diferentes fontes de gordura para vacas em lactação. Por exemplo, Mohamed et al. (1988) não observaram aumento na concentração de cálcio no soro sangüíneo, ao testar a inclusão ou não de grãos de soja sob diferentes formas (10.0; 9.8; 9.9 e 10.3 mg/dL para os animais do grupo controle ou recebendo grãos de soja na forma de óleo livre, grãos inteiros ou grãos tostados, respectivamente). Do mesmo modo, West e Hill (1990) não obtiveram aumento na concentração de cálcio sérico ao adicionarem sais cálcicos à dieta de vacas em lactação (9.49 e 9.64 mg/dL para os animais do grupo controle e com suplementação). Por outro lado, Steele (1984) observou que as vacas em lactação desenvolveram hipocalcemia, quando receberam 2.5 kg/dia de óleo de amendoim ou 2.2 kg/dia de grãos de amendoim moídos. Aumentando o consumo do mineral de 50 para 75 g/dia os valores de cálcio sangüíneo voltaram ao normal em torno de um período de cinco dias de fornecimento. A possibilidade desta ocorrência ser devida à formação de sabões indigestíveis em dietas altas em lipídeos não foi testada no trabalho, uma vez que as perdas fecais não foram determinadas. O autor concluiu que são necessárias mais pesquisas para definir os níveis ótimos de cálcio e de magnésio em dietas com diferentes fontes de gordura e também para explicar o motivo dos animais terem desenvolvido esta doença carencial ao consumirem as dietas.
CONCLUSÕES
A inclusão de gordura na forma de sebo ou gordura protegida ou grãos de soja integral triturados para atingir em torno de 6 % de EE na dieta total de vacas leiteiras na fase inicial da lactação não altera os níveis séricos de glicose, enzima g-glutamil-transferase e cálcio; mas a gordura protegida aumentou os níveis de colesterol, triglicerídeos, ácidos graxos não esterificados e uréia, os grãos de soja aumentaram o nível de colesterol e o sebo aumentou o nível de uréia no soro.
LITERATURA CITADA
© 2004 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la04012t2.jpg] [la04012t1.jpg] |
| |||||||||

{kind=link}
{kind=link}