Archivos Latinoamericanos de Produccion Animal
Asociacion Latinoamericana de Produccion Animal
ISSN: 1022-1301 EISSN: 2075-8359
Vol. 12, Num. 3, 2004, pp. 119-125
Sintitul1
Archivos Latinoamericanos de Produccion Animal, Vol. 12, No. 3,
Septiembre-Deciembre, 2004, pp. 119-125
Alternative animal models to estimate heritabilities and
genetic correlations between direct and maternal effects of Pre and
Post-Weaning weights of Tabapuã cattle1
Modelos animais alternativos para estimar heredabilidade e
correlaçoes geneticas entre efeitos direitos e maternal de pesosas pre e
post desmamedo gado Tabapuã
P. B. Ferraz Filho2 A. de A.
Ramos3, L. O. C. da Silva4, J. C. de
Souza5, M. M. de Alencar4
1The authors thank CAPES for financing this study. 2Send correspondence to P.B. Ferraz Filho, Av. Ranulpho Marques Leal,3484 . Cx.
Postal 210, 79603-011 - Três Lagoas - MS Brazil E-mail:pbferraz@ceul.ufms.br fax 55-67-5093760 3Faculdade de Ciências Agrárias, Universidade Estadual Paulista 4Brazilian Agricultual Research Corporation- (Embrapa) 5Departamento de Zootecnia, Universidade Federal do Paraná
Departamento de Ciências Naturais, Universidade Federal de Mato Grosso do Sul, Brasil
Recibido Mayo 05, 2003.
Aceptado Septiembre 10, 2004
Code Number: la04015
ABSTRACT. The Tabapuã is a Zebu polled breed used for beef production, with white or gray hair
coat and pigmented skin. Weaning (W205, 30,746 observations), twelve-month (W365, 27,701 observations)
and 18-month (W550, 18,493 observations) weights of Tabapuã cattle, raised on pastures in several regions
of Brazil, from 1975 to 1995, were analyzed with four different single trait animal models (including or not
the maternal genetic and/or the permanent environmental effects with the direct genetic effect). Genetic
parameters and covariance components were estimated by restricted maximum likelihood method. For all three
traits the most complete model that included all three random effects was the best. The model that included
only direct genetic effects overestimated the additive genetic variance and heritability (0.27, 0.24 and 0.15 for
W205, W365 and W550, respectively). When the permanent environmental effect was included in the model,
the portion of the total variance due to it was 0.09, 0.06 and 0.02 for W205, W365 and W550, respectively, and
the estimate of the additive genetic variance decreased. The inclusion of the additive maternal effect instead
of the permanent environmental effect did not change the estimate of the additive direct variance.
Heritability estimates, obtained with the most complete model, were low: 0.16, 0.17 and 0.13 for direct genetic effects,
and 0.10, 0.03 and 0.03 for maternal genetic effects, for W205, W365 and W550, respectively. Estimates of
genetic correlation between direct and maternal effects were negative, showing an antagonism between such
effects. Maternal effects still remained evident after weaning, though they became less important.
Keywords: genetic parameters, direct and maternal genetic effects, genetic correlation.
RESUMO. Dados de pesos ao desmame (P205), ao ano (P365) e ao sobreano (P550) de bovinos da
raça Tabapuã, criados em regime de pastagens em diversas regiões do Brasil, no período de 1975 a 1995,
foram estudados em análise univariadas, sob quatro diferentes modelos de análises (incluindo ou não os
efeitos genético materno e de ambiente permanente, além do efeito aditivo direto). Os componentes de
(co)variância e parâmetros genéticos foram estimados pelo método da máxima verossimilhança restrita (REML), usando
o aplicativo MTDFREML Para as três características, o modelo mais completo forneceu o melhor ajuste
dos dados. O modelo que incluiu somente o efeito genético direto do animal superestimou a variância
genética aditiva, inflacionando o valor de herdabilidade
(0,27, 0,24 e 0,15para P205, P365 e P550,
respectivamente). Quando o efeito de ambiente permanente foi incluído no modelo, a variância genética aditiva decresceu e
a porção da variância total em função deste efeito reduziu com o aumento da idade (0,09; 0,06 e 0,02 para
P205, P365 e P550, respectivamente). A inclusão do efeito genético materno em vez do efeito de ambiente
permanente, praticamente não alterou o valor da variância genética aditiva direta. As estimativas de herdabilidade, obtidas sob o modelo mais completo, foram baixas: 0,16; 0,17 e 0,13 para os efeitos direto e 0,10; 0,03 e
0,03 para os efeitos maternos, para P205, P365 e P550, respectivamente e as estimativas de correlações
genéticas entre os efeitos direto e materno foram negativas, evidenciando antagonismo entre estes efeitos. Efeitos
maternos permaneceram ainda evidentes após a desmama, porém menos importantes.
Palavras-chave: parâmetros genéticos, efeitos genéticos direto e materno, correlação genética
INTRODUCTION
In mammals, the dam's contribution to
the phenotype of her progeny is greater than the
sire's, since besides the transmission of the additive
genetic effect to the progeny, the dam provides her
progeny with the maternal environment. Maternal effects
can be defined as any contribution, influence or
impact upon the individual's phenotype, directly
attributed to the dam, which can be caused by genetic
or environmental effects or by a combination of
both. Maternal effects can be manifested in
fertilization, during pregnancy or lactation, and can be
transitory or persist throughout the entire life of the
animal (Hohenboken, 1985). According to Murphey
(1998), it is necessary to consider the maternal effects in
the intrauterine environment during pregnancy and postnatally. These effects include the quality
and quantity of protection provided by the dam
through her behavior, and the passive immunity that
she passes on to her offspring, as well as the
genetic ability for adapting to the environment.
Meyer et al. (1994) considered the quantity
and quality of colostrum and milk that are
directly provided by the dam as the most important
maternal effect. According to Fries and
Albuquerque (1998), besides nursing, the dam is very effective
in determining the macro and microenvironments in which she raises her progeny. Therefore her
decisions when choosing these environments will
directly affect her progeny. The dam also teaches her
progeny to defend itself and to search for water and
food. However, as time goes by this dependency
decreases and terminates at weaning, as maternal bonds
with the offspring are broken.
Thus, besides being influenced by its own
genetic code (direct genetic effect), the initial stages of
an animal's development (mainly until weaning),
are influenced by the dam's genotype for maternal genetic effects and also by the environment,
which acts upon both genotypes (Hohenboken ad
Brinks, 1971).
It is important to quantify maternal influence
and the magnitude of the correlation between the
direct and the maternal genetic effects on the
offspring's body weight in beef cattle in order to
develop effective breeding programs, including
unbiased estimation of heritability for this trait. If
the correlation were positive or null, selection based on weight at weaning normally would not be a
problem. However if the correlation is negative,
genetic progress will be compromised. Part of the
gain obtained by increasing the animal's growth
rate would be accompanied by a reduction of the
dam's maternal ability, and vice-versa (Mercadante
et al., 1995).
Many authors have reported negative
genetic correlations between direct and maternal effects
for several traits and breeds. Therefore, the inclusion
of maternal effect in the models for estimation of
genetic parameters has been suggested (Sousa et
al., 1995, Cobucci et al., 1997), as well as for genetic
evaluations (Reyes et al., 1994).
The Tabapuã is a Zebu polled breed used for
beef production, which has a white or gray hair coat
and pigmented skin. The purpose of this study was
to estimate (co)variance components, heritabilities
and genetic correlations between direct and
maternal effects of pre and post-weaning weights of Tabapuã cattle raised on pasture, in different regions of
Brazil. Estimates obtained with single-trait analyses
with four different models, were compared to quantify
the importance of direct and maternal genetic effects,
as well as maternal permanent environmental effect.
MATERIALS AND METHODS
Weight data were obtained from the
Brazilian Zebu Breeders Association (ABCZ), for a 21
year period (1975 1995) for animals raised on
pasture on farms in several different regions of Brazil.
The information was provided by the National Beef
Cattle Research Center (CNPGC), which belongs to
the Brazilian Agricultural Research Corporation (Embrapa).
Weights were adjusted to standard ages of
205 days (weaning), 365 days (one year) and 550
days (yearling), using the daily gain between the
two weightings closest to each of these standard ages.
Data summary of the three traits studied is
shown in Table 1.
To estimate variance and covariance
components, as well as the genetic parameters for W205,
W365 and W550, four different univariate animal
models were compared for all three traits. These
models contained the random direct genetic effects, with
and without maternal genetic effects and the
maternal permanent environmental effects, as well as the
fixed effects of contemporary group (farm, federation unit, sex, season and year of birth), and age of dam at calving as a covariable (linear and quadratic effects).
Model 1 included the animal's direct additive genetic effect. Model 2 also included the uncorrelated permanent environmental effects. Model 3 included
the animal's direct genetic effects and the dam's maternal genetic effect. Model 4 included all three effects: direct genetic, maternal genetic and permanent
environmental effects.
The most complete model, model 4 in matrix form can be described as:
where,
Y is the vector of observations (W205, W365 and W550); β is the vector of fixed effects in the model, associated with the incidence matrix X; a
is the vector of random direct genetic effects associated with the incidence matrix Z; m is the vector of maternal genetic effects associated with the
incidence matrix M;
pe is the vector of the random maternal permanent environmental effects associated with the incidence matrix W; and e is the vector of the random residual
effects.
The maternal and the direct genetic effects are assumed to be correlated with each other, while the residual and permanent environment effects are not
correlated with each other, nor are they correlated with the genetic effects. The basic structure of variance and covariance matrix for model 4 can be described as:
The estimated components of variance and covariance included: , the phenotypic
variance; , the direct additive genetic variance; , the maternal additive genetic variance; , the variance
due to the dam's permanent environmental effect; , the residual variance; and , the covariance between the direct and the
maternal genetic effects. The following parameters are
derived from the estimates of these components:
= /
= direct heritability, = / = maternal heritability and
= (+ 0,5+1,5) / = total heritability, which according
to Willham (1972) is the regression of the entire genotype of an animal (direct and maternal) on
its phenotype, where = phenotypic variance of the
trait under consideration, given by: . The
correlation between the direct and the maternal genetic effects,
= /(.)1/2, and the proportion of the
permanent environmental effect relative to the total
phenotypic variation, c2 = /, were also computed.
,
where A is the relationship matrix among animals and
I is an identity matrix of appropriate order.
The estimates of the variance and
covariance components and the genetic parameters
were obtained by the derivative free restricted
maximum likelihood method (DFREML), using the MTDFREML (Multiple Trait
Derivative-Free Restricted Maximum Likelihood)
programs developed by Boldman et al. (1993).
The statistical comparison of one model
with another was made using the likelihood ratio
test, which determines the difference between the logarithms of the likelihood functions (log L),
and compares it to a Chi-square value. Deviations
of logarithms of likelihood were obtained from
models that differed in only one random factor.
These deviations were then multiplied by two, the
product of which constituted the l. This l was then
compared with the value from the Chi-square table
(c2) with one degree of freedom at a 95% probability
level, according to the following rule (Dobson, 1990):
l = 2 (log likelihood of model [b] - log
likelihood of model [a])
If l ³ c21,
0.95, model [b] is accepted as fitting the data significantly better than model [a]. If l < c21, 0.95, model [a] and model [b] are not
significantly different, where a is the model with
n parameters and b is the model with n+1
parameters.
Results and Discussion
The estimates of total phenotypic, additive
direct, additive maternal, permanent environmental,
and residual variances, and the covariances
between direct and maternal genetic effects for the weights
at 205, 365 and 550 days of age, as well as log L,
are shown in Tables 2, 4 and 6, respectively.
Estimates of genetic parameters are summarized in Tables 3,
5 and 7, respectively, and values of l, for models comparison, are shown in Table 8.
Table 2. Estimates of (co)variance components
(kg2) for standardized weight at 205 days of age (W205)
Model
Log L
Model 1
436.68
117.83
-
-
318.85
-
-190.72
Model 2
426.97
63.53
-
39.82
323.62
-
-47.57
Model 3
427.77
67.57
64.28
-
323.48
-27.56
-10.21
Model 4
427.23
67.24
42.72
19.02
320.79
-22.54
0
,,,,,= components of phenotypic, additive direct, additive maternal, permanent environmental
and residual variances, and covariance between direct and maternal effects, respectively.
Log L = log likelihood, expressed as a deviation from Model 4.
l = log likelihood, expressed as twice the deviation from Model 4.
Weight at weaning (W205)
Weaning weight was strongly influenced by
maternal genetic and permanent environmental
effects (Models 2, 3 and 4). The estimates of obtained
by Model 1 were biased upward when compared with those obtained by the other models. When
the permanent environmental effect was added
(Model 2), a substantial increase in log L compared to
Model 1 occurred, showing the importance of this effect
on weight at weaning (Table 8). The inclusion of
this effect decreased the estimate of with regard
to Model 1, and was responsible for 9% of the
total variation in W205. In fact, part of the obtained with Model 1 (117.83 kg2) was due to the dam's
effect (Model 2). The inclusion of this effect, resulted in
a decrease of 44.4% for (Table 3).
When W205 was analyzed with Model 3,
which included the maternal genetic effect of the
dam, instead of the permanent environmental effect,
the estimate of increased slightly (Table 2),
resulting in a slight increase in the estimate of direct heritability , when compared to Model 2 (Table 3).
The inclusion of the maternal genetic effect resulted in
a substantial improvement in log L compared to
Model 1, and demonstrated the strong maternal effect
on weight at weaning (Table 8). The direct and
maternal heritability estimates were similar: 0.16 and
0.15, respectively.
Model 4, which included both maternal genetic and permanent environmental effects
was substantially better than model 3 (Table 8). Model
4 reduced the estimate of by partitioning the variance of total dam maternal effects into genetic
and permanent environmental portions.
Direct heritability estimates obtained in this
study for weight at 205 days were low, indicating that
the expected genetic progress by selection for this
trait is not to be high (Table 3).
The estimate of the correlation between direct
and maternal genetic effects was high and negative
(-0.42), which indicates an antagonism between
these effects, making total heritability smaller than
the direct heritability (Table 3).
Weight at one year (W365)
Though relatively less important, the
maternal effects (additive and permanent
environmental) remained evident after weaning (Tables 4 and 5),
and were responsible for 6.6 to 2.7% of the
phenotypic variance of body weight at 365 days of age, and
when included in the model, contributed to a decrease
in direct heritability in models 3 and 4 (Table 5).
Estimates of direct heritability obtained in
this study for weight at 365 days of age, are within
the range of those reported by many authors for
different breeds and methods (Eler et al.,1995; Reyes
et al., 1994; Salles, 1995).
According to the estimates obtained using
Model 3 for W365, and as with W205, it is clear that the maternal heritability () was overestimated,
not including the maternal permanent environmental effect in the model, as is shown by the estimates
with Model 4. Failure to include the maternal
permanent environmental effect in the Model could explain
the estimate of maternal heritability of 0.14 reported
by Mercadante and Lôbo (1997), for the Nelore breed.
Table 3. Estimates of genetic parameters for standardized weight at 205 days of age
Model
Model 1
0.27
-
0.27
-
-
-
0.73
Model 2
0.15
-
0.15
-
-
0.09
0.76
Model 3
0.16
0.15
0.14
-0.42
-0.06
-
0.76
Model 4
0.16
0.10
0.15
-0.42
-0.05
0.04
0.75
, , ,, , and = direct, maternal and total heritabilities, genetic correlation between direct and
maternal effects, covariance between direct and maternal effects as a proportion of the phenotypic variance, variance due to the
permanent environment as a proportion of the total phenotypic variance, and fraction of total phenotypic variance due to the
residual effects, respectively.
Table 4. Estimates of covariance components
(kg2) for standardized weight at 365 days of age
Model
log L
Model 1
725.64
177.52
-
-
548.12
-
-73.83
Model 2
715.46
119.38
-
46.41
549.66
-
-2.99
Model 3
719.25
121.68
47.31
-
557.99
-7.72
-89.78
Model 4
715.90
121.94
19.20
36.43
548.04
-9.71
0
, ,,,,= components of phenotypic, additive direct, additive maternal, permanent environmental
and residual variances, and covariance between direct and maternal effects, respectively.
The estimate of maternal heritability of 0.07
(Table 5), obtained from Model 3, is similar to the
estimate of 0.06 obtained by Robinson and O'Rourke
(1992), for the Brahman breed, and equals that of Magnabosco
et al. (1996). Nevertheless, it is smaller than estimates by ELER
et al. (1995), Reyes et al. (1994) and Salles (1995), for the Nelore breed.
However, when Model 4 was used, the estimate became
equal to 0.03, much smaller than the estimates reported
in the literature.
The estimates of covariance between direct
and maternal genetic effects were negative (-7.72 provided by Model 3 and -9.71 by Model 4),
which are similar to those (-7.79 and -8.99) obtained
by Mercadante and Lôbo (1997), when the same
models were utilized.
The estimates of correlation between direct
and maternal genetic effects for W365 were negative
and small in magnitude: -0.10 and 0.20 with Models
3 and 4 respectively. They were, however, higher
than those obtained by Mercadante and Lôbo (1997).
A possible cause of such negative correlations is
the fact that a negative environmental covariance between the dam and her progeny is not
considered (Baker 1990; Meyer, 1992 and Swalve, 1993).
Total heritability estimates () obtained for
body weight at 365 days of age ranged from 0.16 to
0.24. Model 1 provided the highest value, mainly
because it did not account for maternal effects.
Weight at yearling (W550)
As expected, the influence of the maternal
effect on the estimates was smaller for this trait (Table
6, Model 2), as the offspring becomes less
dependent on its dam (Sousa et al., 1999). The estimate of under Model 2 was less than that obtained
by Model 1. The permanent environmental effect
was responsible for 2.3% of the total variation (Table
7, Model 2).
The estimate of with Model 3, which
considered the maternal genetic effect instead of the
permanent environmental effect, was 5.3% greater than
that obtained with Model 2 (Table 6), increasing the estimate (Table 7). The estimate of was responsible for 3.6% of the total phenotypic
variation (Table 7).
Finally, the analysis with model 4 provided
a higher estimate of than that obtained with
Model 2, and lower than those obtained with Models 3
and 1 (Table 6). The estimate of was 12.6%
smaller than that obtained with Model 3, and was
about 55.3% smaller than that obtained with Model 2.
The covariances between direct and
maternal effects were negative (-21.50 and 29.02, for models 3 and 4, respectively), contributing to a
slight decrease in the total heritabilities when compared
to direct heritabilities. Permanent environmental
effects were low.
The estimates of direct and total heritability for W550 were low, indicating that the expected genetic progress through selection
would not be high.
Table 5. Estimates of genetic parameters for standardized weight at 365 days
Model
Model 1
0.24
-
0.24
-
-
-
0.76
Model 2
0.17
-
0.17
-
-
0.06
0.77
Model 3
0.17
0.07
0.19
-0.10
-0.01
-
0.78
Model 4
0.17
0.03
0.16
-0.20
-0.01
0.05
0.77
, , ,, , and = direct, maternal and total heritabilities, genetic correlation between direct and
maternal effects, covariance between direct and maternal effects as a proportion of the phenotypic variance, variance due to the
permanent environment as a proportion of the total phenotypic variance, and fraction of total phenotypic variance due to the
residual effects, respectively.
Table 6. Estimates of covariance components
(kg2) for standardized weight at 550 days of age
Model
Log L
Model 1
1682.05
256.84
-
-
1425.21
-
-9.64
Model 2
1674.98
216.88
-
38.06
1420.03
-
-3.58
Model 3
1679.37
228.43
54.71
-
1417.73
-21.50
-6.81
Model 4
1675.25
225.06
47.83
17.00
1414.38
-29.02
0
, ,,,,= components of phenotypic, additive direct, additive maternal,
permanent environmental and residual variances, and covariance between direct and maternal effects, respectively.
l = log likelihood, expressed as twice the deviation from the model with the highest value.
Table 7. Genetic parameters for adjusted weight at 550 days of age
Model
1
0.15
-
0.15
-
-
-
0.85
2
0.13
-
0.13
-
-
0.02
0.85
3
0.14
0.04
0.13
-0.19
-0.01
-
0.84
4
0.13
0.03
0.12
-0.28
-0.01
0.01
0.84
, , ,, , and = direct, maternal and total heritabilities, genetic correlation between direct and
maternal effects, covariance between direct and maternal effects as a proportion of the phenotypic variance, variance due to
the permanent environment as a proportion of the total phenotypic variance, and fraction of total phenotypic variance due to
the residual effects, respectively.
Table 8. Differences between l values from
four models used in one-trait analyses for weights at 205 (W205), 365 (W365) and
at 550 (W550) days of age
Models
l
W205
W365
W550
Log L [2] log L [1]
286.26*
141.68*
12.12*
Log L [3] log L [1]
361.04*
327.23*
5.66*
Log L [4] log L [2]
95.16*
5.98*
7.16*
Log L [4] log L [3]
20.42*
179.56*
13.62*
*Significant (P<0.05)
Conclusions
Direct heritability estimates were, in
general, higher than estimates of total heritability. This
means that, when maternal and permanent
environmental effects are not considered, genetic parameter
are overestimated, and consequently the estimates
of breeding values will be unreliable.
Given the low estimates of direct and
total heritability obtained for these herds, information
on relatives should be included in the selection process.
Estimates of the correlation between maternal
and direct genetic effects were negative, showing
an antagonism between these effects.
Literature cited
Baker, R.L. 1990. The role of maternal effects on
the efficiency of selection in beef cattle: a review. Proc.
N. Z. Soc. Anim. Prod., 43:405-419.
Boldman, K.G., L.A. Kriese, L. D. Van Vleck, S.
Kachman. 1993. A Manual for Use of MTDFREML. A set
of programs to obtain estimates of variance and covariance [Draft]. Lincoln, NE U.S. Department
of Agriculture, Agriculture Research Service, 115p.
Cobucci, J. A., A. I. G. Oliveira, T. M. Gonçalves.
1997. Parâmetros genéticos de características
reprodutivas em suínos híbridos - comparação de métodos
usados na estimativa. In: Reunião Anual da Sociedade Brasileira de Zootecnia, 34, 1997, Juiz de Fora,
MG. Anais. v. 3, p. 314-316.
Dobson, A. J. 1990. An introduction to generalized
linear models. Chapman and Hall, Melbourne, 174 p.p.
Eler, J. P., L. D. Van Vleck, J. B. S. Ferraz, R. B. Lôbo.
1995 Estimation of variances due to direct and
maternal effects for growth traits of Nelore cattle. J. Anim.
Sci, 73: 3253-3258.
Fries, L. A, L. G. Albuquerque. 1998 Pressuposições
e restrições dos modelos animais com efeitos
maternos em gado de corte. In: Paranhos da Costa, M. J. R.
and Cromberg, V. U. Ed: Comportamento Materno em Mamíferos (Bases Teóricae e Aplicações
aos Ruminantes Domésticos). São Paulo. Brazilian
Society of Ethology (Sociedade Brasileira de Etologia). P.
161-178.
Hohenboken, W. D. 1985 Maternal Effects. In: General
and quantitative genetics. Amsterdam: Elsevier
Science, 1985. p. 135-149.
Hohenboken, W.D., J. S. Brinks.
1971. Relationships between direct and maternal effects on growth
in Herefords: Partitioning of covariance between
relatives. J. Anim. Sci. 32:26-34.
Magnabosco, C de U., T. R. Famula, R. B. Lôbo, A. de
los Reyes, M. Ojala. 1996. Estimativas de
parâmetros genéticos e de ambiente de características
de crescimento em bovinos da raça Nelore. In:
Reunião Anual da Sociedade Brasileira de Zootecnia, 33,
Fortaleza. Anais. SBZ, p.142-144.
Mercadante, M. E. Z., R. B. Lôbo, A. R. Borja.
1995. Parámetros genéticos para características de
crecimiento en cebuínos de carne. Arch. Latinoam. Prod. Anim. 3: 45-89.
Mercadante, M.E.Z., R. B. Lobo. 1997. Estimativas de
(Co) variâncias e parâmetros genéticos dos efeitos direto
e materno de características de crescimento de
fêmeas de um rebanho Nelore. R. Bras. Zootec. 26: 1124-1133.
Meyer, K. 1992 Variance components due to direct
and maternal effects for growth traits of Australian
beef cattle. Livest. Prod. Sci., 179-204.
Meyer, K., M. J. Carrick, B. J. P. Donnley. 1994.
Genetic parameters for milk production of Australian beef
cows and weaning weight of their calves. J. Animal Sci., 72:1155-1165.
Murphey, R. M. 1998. Análise genética do
comportamento animal. In: Paranhos da Costa, M. J. R. e Cromberg,
V. U. Ed: Comportamento Materno em Mamíferos
(Bases Teóricae e Aplicações aos Ruminantes Domésticos).
São Paulo. Brazilian Society of Ethology. p. 161-178.
Reyes, A. de los, R. B. Lôbo, H. N. Oliveira, S. P. F.
Bezerra, Jr. R. P. L. Gestal. 1994. Estimativa de efeitos
genético direto e maternal para pesos até 365 dias, usando
modelo animal bivariado em rebanhos Nelore. In
Reunião Anual da Sociedade Brasileira de Zootecnia,
31, Maringá, PR. Anais..., p. 167.
Robinson, D.L., P. K. O'Rourke. 1992. Genetic
parameters for liveweights of beef cattle in the tropics. Aust.
J. Agric. Res. 43:1297-1305.
Salles, P. de A. 1995. Critérios de seleção para
características de crescimento em machos da raça
Nelore. Ribeirão Preto, SP, 1995. Dissertação (Mestrado
em Ciências) Universidade de São Paulo, 1995.
Sousa, W. H., C. S. Pereira, J. A. G. Bergmann, F. L.
R. Silva. 1999. Estimativas de Componentes de (Co) variância e herdabilidade Direta e Materna de
Pesos Corporais em Ovinos da Raça Santa Inês. R. Bras.
de Zootec., 28(6):1252-1262.
Sousa, W. H., R. A. Torres, P. R. M. Leite. 1995.
Estimativas de componentes de variância devido aos efeitos
direto e materno para características de crescimento
em caprinos. In: Reunião Anual da Sociedade
Brasileira de Zootecnia, 32, Brasília, DF. Anais.... p. 721-722.
Swalve, H. H. 1993. Estimation of direct and
maternal (co)variance components for growth traits in
Australian Simental beef cattle. J. Anim. Sci., 110:241-248.
Willham, R. L. 1972. The role of maternal effects in
animal breeding: III. Biometrical aspects of
maternal effects in animals. J. Anim. Sci., 35(6): 1288-1293.


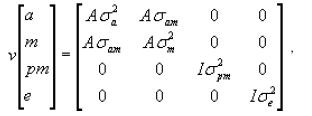
 ,
,