 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 12, 4 Suppl. 1, Deciembre, 2004, pp. 7-11 Estudio de las relaciones morfológicas existentes en las abejas domésticas (Apis mellifera L) de las Islas Canarias Study of the morphological relationship existent in the CanaryIsland honeybee (Apis mellifera L.) F. Padilla Álvarez[1] y R. Hernández Fernández Departamento de Zoología, Universidad de
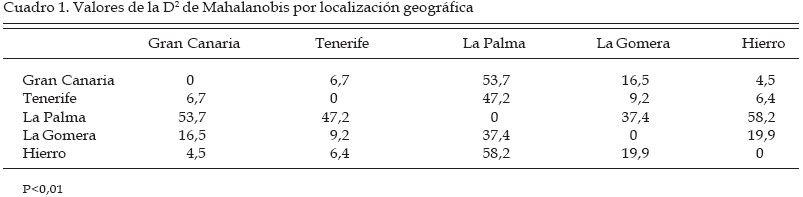
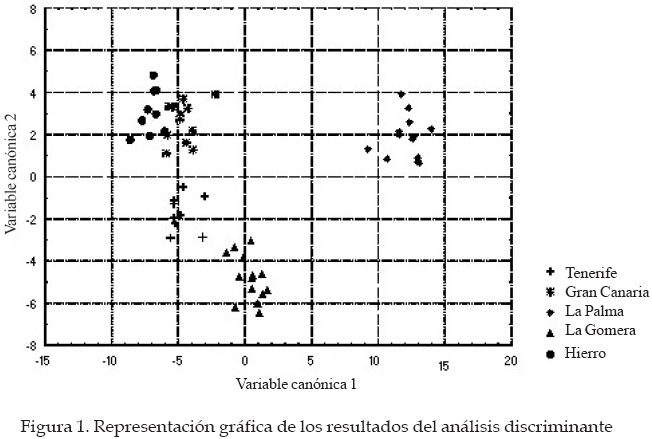
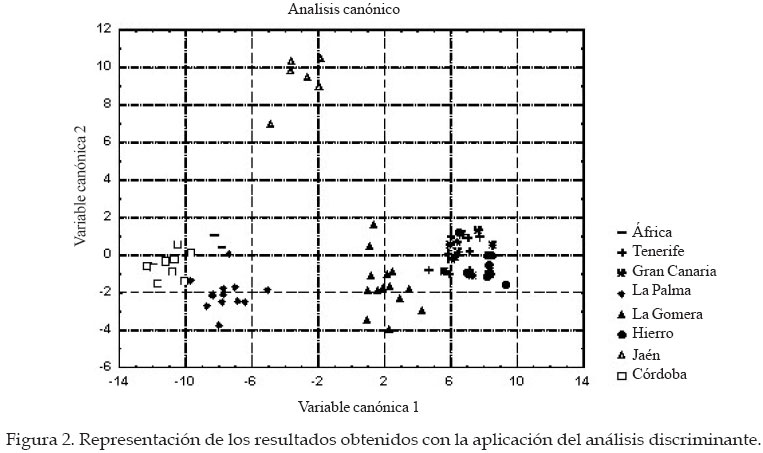
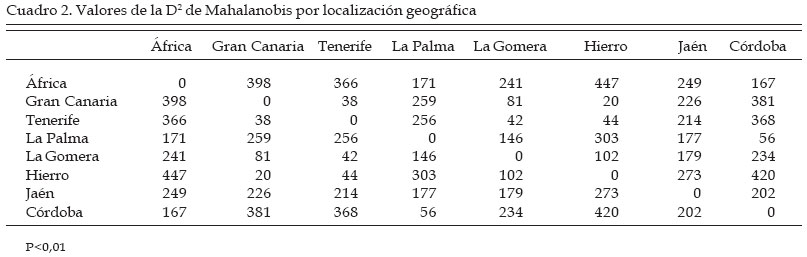
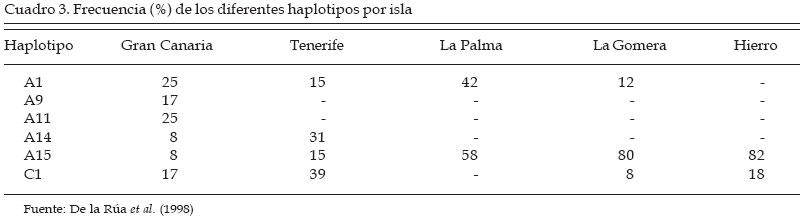
Córdoba Code Number: la04018 ABSTRACT The genetic origin of the honey bees (Apis mellifera) that lives in the CanaryIsland is ignored. It has been speculated with the possibility that the Guanches, first island inhabitant with African origin, brought with them native bees from the continent. It is known that the first Spanish colonist transport beehives from the Iberian Peninsula, and the quality of the canary honey allowed that in the XVI century it was exported in big quantities toward Europe. This paper evaluates morphological traits in samples of honeybee workers coming from apiaries located in five islands of the archipelago. A total of 27 morphological variables was taken of each insect. The discriminant analyses carried out, using as classificatory variable the beehive of origin of the work bees, indicate that significant differences exist among the groups. Four morphological groups were identified. One of them includes the bees coming from Gran Canaria and Hierro, between this group and the one formed by the workers coming from La Gomera is located the one from Tenerife, and the one from La Palma forms an independent group. The possible relationship of these bees was estimated with another discriminant analysis that included data coming from samples of the south of Spain (A. mellifera iberica) and north of Africa (A. mellifera intermissa). Key words: Order Hymenoptera, Family Apidae, Apis mellifera, Biogeography, Morphometry, CanaryIsland. RESUMEN Se desconoce el origen genético de las abejas domésticas (Apis mellifera L.) que pueblan el Archipiélago Canario. Se ha especulado con la posibilidad de que los Guanches, primeros pobladores de las islas de origen africano, llevaran consigo abejas oriundas desde el continente. Sabemos que los primeros colonizadores españoles sí llevaron colmenas desde la Península Ibérica, y la calidad de la miel canaria permitió que en el siglo XVI se exportara en grandes cantidades hacia Europa. El presente trabajo evalúa características morfológicas en muestras de abejas obreras procedentes de colmenares situados en cinco islas del archipiélago. De cada insecto se tomaron valores de 27 variables morfológicas. Los análisis discriminantes realizados, empleando como variable clasificatoria la colmena de procedencia de las abejas obreras, nos indican que existen diferencias significativas entre grupos. La representación gráfica de los resultados del análisis canónico muestra un agrupamiento de las muestras procedentes de Gran Canaria y Hierro, entre este grupo y el formado por las muestras de La Gomera se sitúan las procedentes de Tenerife; las abejas de La Palma forman un grupo independiente. Para poder especular sobre el posible parentesco de estas abejas, realizamos otro análisis discriminante incluyendo los datos procedentes de muestras del sur de España (A. mellifera iberica) y norte de África (A. mellifera intermissa). Palabras claves: Orden Himenoptera, Familia Apidae, Apis mellifera, Biogeografía, Morfometría, Archipiélago Canario. Introducción Se desconoce el origen de las abejas domésticas (Apis mellifera) que pueblan el Archipiélago Canario. Se ha especulado sobre la posibilidad de que los Guanches, primeros pobladores de las islas, las introdujesen desde el continente africano. Esta hipótesis es apoyada por las referencias que hacen de la miel canaria algunos autores antiguos, como el poeta latino Quinto Horacio Flaco. Es conocido de forma más cierta que los primeros colonizadores españoles llevaron colmenas desde la Península Ibérica. La calidad de la miel producida en las islas permitió que en el siglo XVI se exportara en grandes cantidades hacia Europa (Crosby, 1988). Actualmente en el Archipiélago Canario hay entre 18.000 y 20.000 colmenas que producen unas 220 de miel, consumida en el mercado interior. Hasta el momento, con la excepción del trabajo realizado por Ruttner (1975), sólo Padilla et al., (1997, 98, y 2001a[Comment1] ) han reportado resultados de estudios morfológicos de estos insectos. En el presente trabajo queremos estudiar si las abejas oriundas de estas islas constituyen un grupo morfológico aislado, o bien forman grupos morfológicos independientes más o menos emparentados con las abejas procedentes de la Península Ibérica (A. mellifera iberica) o el norte de África (A. mellifera intermissa) Materiales y Métodos Un total de 58 muestras de abejas obreras de edad desconocida y tomadas de los panales centrales de las colmenas, fueron conservadas en alcohol de 70º hasta su procesado. Las muestras se corresponden con las localizaciones siguientes: Gran Canaria (12), Tenerife (9), La Palma (14), La Gomera (14) y Hierro (9). De cada muestra se diseccionaron entre 25 y 44 abejas siguiendo la técnica descrita previamente por Padilla et al. (2001). Un equipo de análisis de imágenes y el programa IMAGO desarrollado por el grupo de trabajo SIVA de la Universidad de Córdoba fue usado para realizar las mediciones. De cada insecto diseccionado se midieron las variables siguientes: anchura (1) y longitud del ala anterior (2), anchura (3) y longitud del ala posterior (4), longitud de la probóscide (5), longitud del fémur (6), longitud de la tibia (7), longitud del metatarso (8), anchura del metatarso (9), diámetro longitudinal del terguito 2º del gaster (10), diámetro longitudinal del terguito 3º del gaster (11), longitud del esternito 2º del gaster (12), longitud del esternito 3º del gaster (13), longitud del espejuelo de la cera del 3º esternito del gaster (14), anchura del espejuelo de la cera del 3º esternito del gaster (15), distancia entre los espejuelos de la cera del 3º esternito del gaster (16), ángulos del ala anterior A4 (17), B4 (18), D7 (19), E9 (20), G18 (21), J10 (22), J16 (23), K19 (24), L13 (25), N23 (26), O26 (27). Los datos obtenidos fueron estudiados estadísticamente mediante el paquete de programas estadísticos Statistica versión cinco. Básicamente se utilizaron los procedimientos incluidos en el programa “Discriminant Analisys”. Resultados y Discusión El uso de un gran número de características morfológicas mejora la caracterización de los animales integrantes de un grupo morfológico o de una especie, cuanto mayor sea el número de variables empleadas mayor es la riqueza potencial de información taxonómica contenida en las mismas. Por este motivo fue usado un amplio conjunto de variables, para poder caracterizar de una forma adecuada las poblaciones objeto del presente estudio. Además y con el objetivo de reducir la variabilidad existente dentro de cada colonia, debido principalmente al diferente grado de parentesco que presentan las abejas obreras de un mismo enjambre, se usó el valor de las medias para cada característica y colonia estudiada. Desde un punto de vista morfológico las muestras procedentes de diferentes colmenas y localizaciones geográficas son muy parecidas entre si, por lo que no es conveniente establecer diferencias basadas en el color o el tamaño de los insectos. Comenzamos el estudio estadístico realizando un análisis discriminante según el procedimiento “Forward stepwise” incluido en el “Discriminant analisys”, utilizando como variable clasificatoria la isla de procedencia. Los resultados obtenidos muestran la existencia de diferencias significativas entre todas las colonias.Utilizando un valor de 1 para el estadístico “F” de Fischer, el procedimiento elimina de forma automática las variables: 9, 10, 13, 14, 17 y 25. La matriz de clasificación muestra que el 100% de las observaciones se clasifica correctamente en su grupo de origen. Si consideramos que cada isla constituye un grupo biométrico y que dichos grupos se encuentran relacionados, podemos calcular los valores de las D2 de Mahalanobis para poder indagar sobre sus relaciones. Los resultados se muestran en el Cuadro 1 e indican que los grupos de Gran Canaria, Hierro y Tenerife se encuentran muy cercanos, a mayor distancia se sitúa La Gomera y el menos relacionado con los demás es el grupo de La Palma. Los resultados del análisis canónico (procedimiento incluido en el “Discriminant Analisys”) en el que las variables originales son transformadas en variables canónicas se dan en la Figuras 1 y 2. La representación gráfica de los resultados, empleando las dos primeras variables canónicas, que acumulan el 92 % de la varianza, se muestra en la Figura 1. En dicho gráfico observamos la formación de cuatro grupos morfológicos, uno está constituido por las abejas procedentes de las islas de Gran Canaria y Hierro, entre dicho grupo y el formado por las muestras de La Gomera se sitúan las colonias de Tenerife, y las de La Palma constituyen un grupo independiente alejado del resto. Llama la atención el hecho de que todos los datos se incluyan en sus grupos geográficos originales, debido a que las islas no son ecosistemas homogéneos. En términos generales la orografía y la climatología varían de una isla a otra, además dentro de cada isla la variabilidad ambiental también es elevada y las condiciones meteorológicas cambian con los puntos cardinales y la altitud. Frente a esta variabilidad ambiental, los resultados pueden indicar una buena adaptación de los insectos a su entorno, sobre todo en los casos de las islas de La Gomera, La Palma y Hierro. En cada una de estas islas se produce una cierta trashumancia estacional y obviamente transacciones de compra-venta de colonias, pero según los apicultores no es corriente que se importen abejas de otras localizaciones. El posible parentesco de estas abejas fue estimado a través de una serie de análisis discriminantes en los que fueron incluidos datos procedentes de colmenas del norte de África (dos colonias procedentes de Tetuan, A. mellifera intermissa) y del sur de España (seis naturales de Jaén y 12 de Córdoba, A. mellifera iberica). El análisis discriminante siguiendo el procedimiento “Forward stepwise” (aceptando un valor de 1 para el estadístico “F”) incluye en el modelo 24 y elimina las variables: 1, 18 y 19. La matriz de clasificación muestra que el 100% de las colmenas se clasifican correctamente en su grupo de origen. Los estimados de D2 de Mahalanobis (Cuadro 2) difieren significativamente de cero (P < 0,01), lo cual es indicativo de diferencias estadísticamente significativas entre los grupos comparados. Las dos primeras variables canónicas acumulan el 80% de la varianza, y en la representación gráfica plasmada en la Figura 2 se observan cuatro grupos morfológicos: uno integrado por las muestras oriundas de Gran Canaria, Tenerife y Hierro; las procedentes de La Gomera forman un grupo independiente situado entre el anterior y otro formado por las abejas de La Palma, Córdoba y África; las colmenas de Jaén establecen un grupo independiente y bastante alejado de los otros. De la Rúa et al. (1998) estudian el ADN mitocondrial de abejas obreras procedentes de 79 colonias oriundas del Archipiélago Canario. Los autores encuentran en la región intergénica tRNA Aleu-citocromo oxidasa II cinco haplotipos de la línea africana (A), uno de la línea de oeste europeo (C) y dos nuevos (denominados como A14 y A15). La frecuencia de estos haplotipos dentro de cada isla se plasma en el Cuadro 3. Al comparar esos resultados con los obtenidos en este estudio se producen algunas curiosidades. Las abejas procedentes de la isla de Hierro se sitúan en el mismo grupo morfológico que las de Gran Canaria y Tenerife. En las muestras de insectos de Gran Canaria se han descrito 6 haplotipos diferentes (A1, A9, A11, A14, A15, C1), en los de Tenerife 4 (A1, A14, A15 y C1), y en Hierro el 82% de los haplotipos encontrados son del tipo A15 (nuevo y sólo descrito en este archipiélago), pero este haplotipo sólo representa el 8% de la población en la isla de Gran Canaria y el 15% en la de Tenerife. De la Rúa en su trabajo plantea la posibilidad de que el haplotipo A15 se halla podido originar a partir del A11 (presente en el sur de la Península Ibérica y ausente en el norte de África), por lo tanto estas abejas estarían emparentadas con las españolas descritas tradicionalmente como pertenecientes a la raza A. mellifera iberica. Además el 18% de los ejemplares de la isla de Hierro estudiados presentan el haplotipo C1 que se corresponde con la línea C de abejas del este de Europa (en este grupo se incluye entre otras la raza A. mellifera ligustica). Es decir, independientemente de la opinión de los apicultores, en esta isla se han introducido abejas de un origen geográfico bastante lejano. Análisis progresivos de resultados parecen conducir hacia diferentes opciones. ¿Qué camino seguir?, máxime cuando los datos procedentes de la aplicación de las técnicas morfológicas clásicas, no se ajustan o son bastante opuestos a los obtenidos con las nuevas técnicas de estudio del ADN (ej. ADN mitocondrial). Los genes implicados en la morfología corporal están sometidos a una selección natural determinada por el ambiente en el que viven las poblaciones, pero probablemente la interacción ADN-mitocondrial /ADN-nuclear no se encuentre sometida a la misma presión selectiva. Agradecimientos Agradecemos la colaboración de la Consejería de Agricultura y Alimentación del Gobierno de Canarias, así como de la Agrupación de Defensa Sanitaria de la isla de La Palma. Literatura Citada
[Comment1]a hay dos referencia con igual cita, sugerimos diferenciarlas con a y b. © 2004 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la04018t3.jpg] [la04018f2.jpg] [la04018t1.jpg] [la04018f1.jpg] [la04018t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}