 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 12, Suppl. 1, Deciembre, 2004, pp. 45-48 Empleo de técnicas moleculares en el apoyo a la conservación del cerdo Chato Murciano The use of Molecular Techniques in the Support to the Conservation of the Shato Murciano Breed of Pig. J. L. Vega-Pla[1], A. M. Martínez[2], B. Peinado, A. Poto2, y J. V. Delgado[3] Ministerio
de Defensa, Laboratorio de
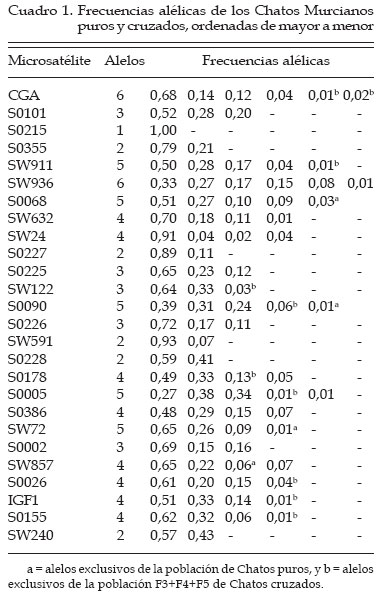
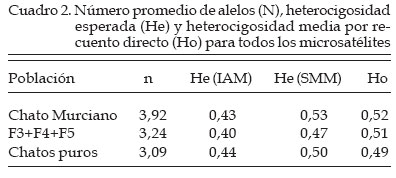
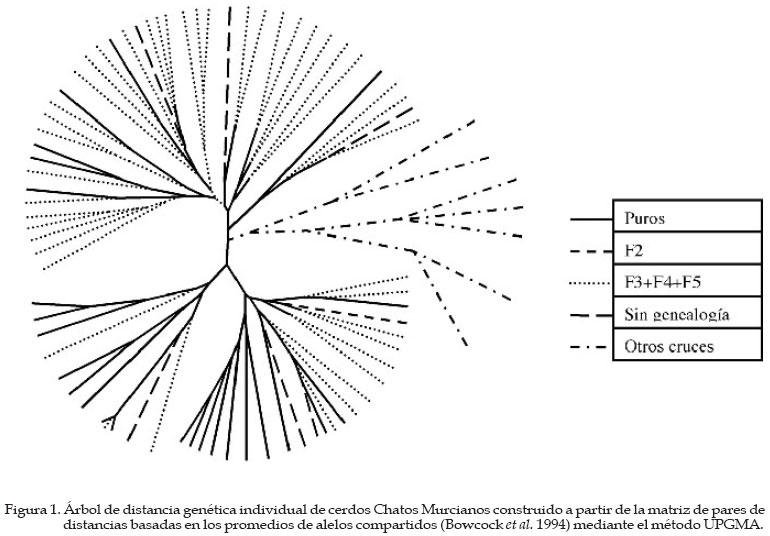
Genética Molecular. Córdoba, España Code Number: la04026 ABSTRACT The use of molecular allelic typing tools in small populations near extinction is evaluated. 83 samples of Chato Murciano pig are typed with 26 microsatellites recommended for biodiversity studies. Allelic frequencies are obtained by direct count. Observed, unbiased and expected heterocigosity and pairs of individual distances based in shared alleles are calculated. An individual tree is built from matrix of distances. Allele number is smaller than other pig breeds. The process of breed conservation can be followed by mean of individual phylogenetic tree drawing. Observing, as even it is necesary the creation of a line by using of a LargeWhite female, is shown an absorption of the genotype of this animal in favour of the Chato Murciano genotypes by generations. This approach also facilitates the selection of the best mating. Key words. Pig, Microsatellites, Genetic distance, Genetic resources RESUMEN Se investiga la aplicación de técnicas de tipificación alélica de marcadores moleculares, para el apoyo de planes de recuperación de poblaciones en vías de extinción. Se recogen muestras de 83 cerdos de raza Chato Murciano y se caracterizan 26 microsatélites seleccionados y recomendados para estudios de diversidad genética en cerdos. Las frecuencias alélicas se determinan por recuento directo. Se calculan las heterocigosidades observada, insesgada y esperada para cada marcador y los pares de distancias genéticas entre individuos basadas en la proporción de alelos compartidos. Con los valores de las distancias obtenidos se construye un árbol filogenético. El número de alelos presentes es claramente inferior al hallado en otras razas porcinas. El proceso de conservación de la raza se puede seguir mediante la construcción de un árbol filogenético individual observando cómo, a pesar de ser necesario crear una línea mediante un cruce por absorción empleando una hembra de raza Large-White, se aprecia una tendencia a la absorción del genotipo de este animal en favor de los genotipos de los Chatos Murcianos a través de las generaciones. Este abordaje facilita también la selección de los cruzamientos más adecuados a partir de los genotipos caracterizados. Palabras Claves: Porcino, Microsatélites, Distancia genética, Recursos genéticos. Introducción El cerdo Chato Murciano tiene su origen en los antiguos cerdos ibéricos del área mediterránea. Durante el primer cuarto del siglo XX fue cruzado con ejemplares de las razas Berkshire, Tamworth y otras razas europeas para mejorar sus caracteres productivos (Martínez et al., 1998). En la última década de este mismo siglo, cuando estaba prácticamente extinto, aparece un gran interés a nivel institucional para la conservación de este cerdo tan característico del este de la península ibérica. Se trata de una raza que se desarrolla en régimen semi-extensivo y su expansión se realiza a partir de tan sólo 14 ejemplares. Después de cinco años de trabajo en la recuperación de esta raza se plantea un estudio genético para evaluar los resultados obtenidos y elaborar futuros proyectos. El objetivo de este trabajo es obtener una información genética objetiva de la población de Chatos Murcianos que permita contribuir al plan de recuperación de esta raza. Para ello se caracteriza la población con una batería de microsatélites, se obtendrán parámetros estadísticos descriptivos de la situación de variabilidad genética y construir un árbol de distancias individuales. Materiales y Métodos Se analiza una muestra de 83 ejemplares de cerdo Chato Murciano, 24 originales y el resto pertenecientes a las generaciones F2 (1), F3 (15), F4 (17), F5 (12) obtenidas después de un cruzamiento inicial con una hembra de la raza Large-White y sementales de la población original. Además se analizan sieteejemplares de Chato Murciano de los que no se conoce su genealogía y otros siete ejemplares resultantes de cruces de Chato Murciano con otras razas. Estos últimos sirven de referencia en el árbol individual de distancias. Los microsatélites seleccionados son los recomendados para los estudios de variabilidad por el grupo de expertos en temas porcinos de la FAO: CGA, S0101, S0215, S0355, SW911, SW936, S0068, SW632, SW24, S0225, SW122, S0090, S0226, S0227, SW951, S0228, S0178, S0005, S0386, SW72, S0002, SW857, S0026, IGF1, S0155, SW240. Se extrae DNA de muestras de pelo, se amplifican mediante PCR las secuencias de interés utilizando cebadores marcados con fluorocromos combinando diferentes marcadores (Martínez et al., 1998). La caracterización de las variantes alélicas se realiza mediante electroforesis de los productos de la amplificación en un secuenciador automático ABI 373 Stretch (Applied Biosystems, Foster City, CA, USA) empleando las aplicaciones GENESCAN" 3.1.2 y GENOTYPER" 2.5 (Applied Biosystems, Foster City, CA, USA). Se calculan las frecuencias alélicas por recuento directo y se determinan la heterocigosidad observada y esperada teniendo en cuenta los modelos de mutación IAM (infinity allele model) o SMM (stepwise mutation model). Para estos cálculos se emplea la aplicación BOTTLENECK (Cornuet & Luikart, 1996). Mediante la aplicación MICROSAT v.1.5b (Minch, 1998) se calculan las distancias genéticas entre individuos basadas en la proporción de alelos compartidos (Bowcock et al., 1994). Con estas distancias se construye un árbol individual filogenético basado en el algoritmo UPGMA (unweighted pair-group method with arithmetic mean) (Sneath & Sokal, 1973) empleando para ello el módulo NEIGHBOR de la aplicación PHYLIP v.3.57c (Felsestein, 1995). Resultados y Discusión El estudio genético de poblaciones lleva a plantearse un gran número de preguntas de las que algunas son contestadas mediante la observación directa del grupo en cuestión, y el resto intenta solucionarse mediante la investigación de caracteres más objetivos como los marcadores genéticos. Recientemente los microsatélites se han configurado como los marcadores de elección debido a algunas de sus ventajas frente a los demás como son que se encuentran en gran número, distribuidos de forma homogénea a lo largo del genoma y presentando un gran nivel de polimorfismo (Edwards et al., 1992). Su estudio parece ser muy interesante en aquellas poblaciones que por su situación dramática de peligro de extinción son más urgentes de caracterizar, de forma que la variabilidad genética y su entidad queden salvadas. Así, está indicado este tipo de estudios de conservación genética y está apoyado enfáticamente por la FAO desde 1993 (FAO, 1998). Los trabajos que en este sentido se han ido realizando, son numerosos y la efectividad y potencialidad informativa de estos marcadores moleculares, está ya comprobada (ej.: Arranz et al., 1996; MacHugh et al., 1998; Martínez et al., 2000a). La raza se encuentra en un proceso de recuperación a partir de muy pocos individuos, hay una consanguinidad grande y una variabilidad genética pequeña. El número de alelos observado es claramente inferior al encontrado en otras razas porcinas (Martínez et al., 2000a; Laval et al., 2000; Ollivier et al., 2001). Algunos alelos se encuentran sólo en los animales originales mientras que hay otros presentes solamente en los animales de la población F3+F4+F5 (Cuadro 1). Es de destacar que los alelos presentes sólo en una u otra población suelen ser aquellos que están presentes con las frecuencias más bajas. El microsatélite S0215 ha resultado monomórfico en todos los animales analizados, y el S0951 es monomórfico en la población de Chatos Murcianos puros. La heterocigosidad observada para el total de los animales analizados (Cuadro 2) presenta valores inferiores a los encontrados en otras razas porcinas (Li et al., 2000), aunque son similares a los encontrados en otras razas minoritarias (Martínez et al., 2000a; Laval et al., 2000; Ollivier et al., 2001). La heterocigosidad observada en los tres casos es claramente superior a la esperada si se considera que el modelo seguido por estos microsatélites es el IAM, sin embargo cuando se tiene en cuenta el SMM, la heterocigosidad observada sólo es claramente superior a la esperada en el caso de la población F3+F4+F5 (Cuadro 2). El modelo que más se ajusta a los resultados esperados es el IAM, teniendo en cuenta las características de la población, como ya afirmaban Shriver et al. (1995) que los microsatélites en los que la unidad repetitiva es de 3 a 5 pares de bases se ajustan mejor al SMM y los microsatélites mono o dinucleotídicos, como en este caso, se ajustan mejor al IAM Se realiza un estudio de distancia genética individual para evaluar la tendencia de los animales a agruparse juntos por poblaciones basado en la proporción de alelos compartidos (Bowcock et al., 1994). Los microsatélites, tienen un grado relativamente alto de heterocigosidad y muchos alelos para cada locus, por lo que cada individuo puede ser definido de una forma casi inequívoca. Esto implica que pueden obtenerse resultados significativos de alelos compartidos entre individuos miembros de las poblaciones estudiadas y que pueden construirse árboles filogenéticos usando los individuos como unidades taxonómicas. El árbol de distancia genética (Figura 1) ha generado tres grupos principales de individuos que comparten características similares, a pesar de que el conjunto de los animales están muy emparentados. Una de las conclusiones que se podría deducir a partir de este análisis es que las F3, F4 y F5 son muy parecidas entre si y a la vez a los animales puros por lo que el efecto del cruzamiento con el ejemplar de Large-White parece que está siendo absorbido. El análisis genético realizado con la batería de microsatélites da como resultado que esta raza presenta un grado de variabilidad genética bajo aunque la heterocigosidad es aceptable, lo que indica que los esfuerzos realizados para su recuperación están teniendo su fruto. El establecimiento de una línea dentro de la raza mediante un cruzamiento con otra raza y una posterior absorción en sucesivas generaciones no está desviando el perfil genético de la población original por lo que se pueden esperar los beneficios que este tipo de estrategia de conservación puedan aportar en un futuro. Finalmente, el estudio de marcadores genéticos como los microsatélites aporta una información muy valiosa y útil en el diseño y seguimiento de un proceso de recuperación de una raza en peligro de extinción. Literatura Citada
© 2004 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la04026t2.jpg] [la04026t1.jpg] [la04026f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}