 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
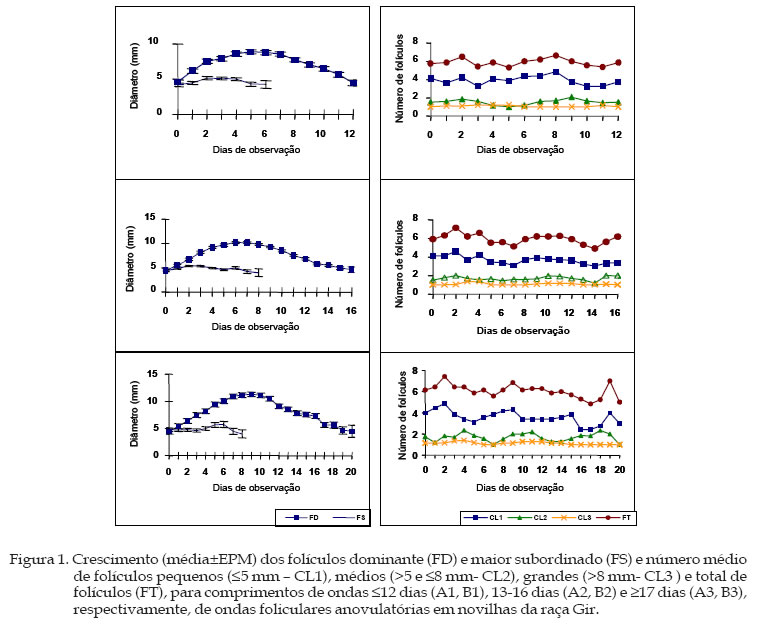
Archivos Latinoamericanos de Produccion Animal, Vol. 13, No. 2, Mayo-Agosto, 2005, pp. 51-55 Dinâmica folicular por ultra-sonografía em novilhas pré-púberes da raça Gir Study of follicular dynamic by real-time ultrasound inprepuberal Gir heifers AR. Reis e Silva, A. de los Reyes1+, M.L. Gambarini, R. Rumpf2, C.C. Oliveira3, B. Dias de Oliveira Filho1 Universidad Federal de Goiás. Departamento de Produção
Animal, Escola de Veterinária,
Goiânia, Goiás, Brasil. Recibido Julio 19, 2004. Code Number: la05007 ABSTRACT. The objective of this research was contribute to knowledge of reproduction physiology in the Gir breed, by ultrasound study of follicular dynamic in prepubertal heifers. Follicular growth was monitored in 13 heifers during 42 consecutively days. A process of successive anovulatory follicular waves was identiffied, with each wave showing a dominant follicle and a variable number of other smaller in size (subordinates). The waves were variables in length, for purposes of analysis, they were classified in three categories as follows: waves of class 1 length (CO1: ≤12 days), class 2 (CO2: 13-16 days) and class 3 (CO3: ≥17 days). Forty-nine waves were thus analyzed: 6 CO1 (12.2%), 31 CO2 (63.3%) and 12 CO3 (24.5%). Differences among length of waves were observed, even between those from the same female. In the CO1 class the mean maximum diameter of the dominant follicle was (P<0.05) shorter (9.4±0.3 mm) than CO2 (10.4±0.2 mm) and CO3 (11.6±0.4 mm). The plateau and rates of growing and atresia of follicles were not different (P>0.05) between CO classes. Similar characteristics on subordinate follicles were not different (P>0.05) among CO classes. Key words: Follicular waves, Gir breed, Reproduction, Zebu cattle RESUMO. O presente trabalho visa contribuir para o conhecimento da fisiologia reprodutiva na raça Gir, através do estudo do desenvolvimento folicular em novilhas. O desenvolvimento folicular foi monitorado, por ultra-sonografia, a cada 24 horas, em 13 fêmeas pré-púberes durante 42 dias consecutivos, e caracterizou-se pela presença de sucessivas ondas de crescimento folicular (OCF) anovulatórias, com cada onda apresentando um folículo dominante e um variável número de outros menores (subordinados). Estas ondas apresentaram-se variáveis em comprimento, por esta razão, foram agrupadas em três categorias: comprimento de onda um (CO1: ≤12 dias), comprimento dois (CO2: 13-16 dias) e comprimento três (CO3: ≥17 dias), para serem melhor analisadas. Foram acompanhadas 49 ondas, sendo 6 CO1, 12,2%; 31 CO2, 63.3% e 12 CO3, 24.5%; ocorrendo ondas de diferentes comprimentos em um mesmo animal. Nas ondas CO1, o diâmetro máximo do folículo dominante foi menor (P<0,05) (9,4±0,3 mm) que para CO2 (10,4±0,2 mm) e CO3 (11.6±0.4 mm). O platô e as taxas de crescimento e atresia, entre os três CO não apresentaram diferenças (P>0,05). Para os folículos subordinados, as características testadas não diferiram (P>0,05) entre as três classes de CO. Palavras-chave: Dinâmica folicular, Gir, Reprodução, Zebu. Introdução A raça Gir, devido a sua rusticidade e resistência ao estresse climático, teve seu desenvolvimento e multiplicação favorecidos nas condições bioclimáticas brasileiras, sendo utilizada para a produção de leite e carne (Ledic, 1994), bem como serviu de base para cruzamentos visando a produção de genótipos leiteiros, por exemplo, o Girolando (Menezes, 1999). A utilização da ultra-sonografia na reprodução de grandes animais possibilitou o entendimento do desenvolvimento folicular, o que permite verificar as particularidades da fisiologia reprodutiva e assim, propor protocolos adequados para a melhoria da eficiência reprodutiva dos bovinos. Este conhecimento, através do entendimento do comportamento das ondas de crescimento folicular (dinâmica folicular), favorece a execução de programas de superovulação (Amstrong, 1993; Adams, 1994) e sincronização do cio (Viana et al., 1997) e da ovulação (Gambini et al., 1998). O desenvolvimento folicular tem sido estudado principalmente em fêmeas de genótipos taurinos (Matton et al., 1981; Savio et al., 1988; Sirois e Fortune, 1988; Ginther et al., 1991; Adams et al., 1994; Evans et al., 1994). Porém, são escassos os estudos deste processo em animais zebuínos (Rhodes et al 1995, Figueiredo et al 1997), especialmente da raça Gir (Gambini et al., 1998). Os animais desta raça têm apresentado resultados insatisfatórios nos protocolos clássicos utilizados para superovulação em programas de transferência de embriões (Freitas et al., 1998). Neste contexto, torna-se importante analisar não só este processo na fêmea adulta, mas acompanhá-lo em animais jovens. Pois, com maior entendimento da fisiologia ovariana poder-se-á manipulá-la mais eficientemente (Adams, 1994; Thatcher et al., 1996).Desta forma, o presente trabalho visa contribuir para o conhecimento da fisiologia reprodutiva na raça Gir, através do estudo da dinâmica folicular em novilhas pré-púberes, nas condições ambientais e sistema de produção extensivo da região Centro-Oeste do Brasil, onde predomina a vegetação de cerrados. Materiais e MétodosO estudo foi desenvolvido no Centro Nacional de Pesquisa em Arroz e Feijão da Empresa Brasileira de Pesquisa Agropecuária (CNPAF-EMBRAPA), Santo Antônio de Goiás-GO, Brasil, de abril a setembro de 2000. Foram utilizadas 13 novilhas da raça Gir, com peso e idade médios no início do experimento de 252±25 kg e 22±2 meses, respectivamente. Os animais escolhidos apresentavam escore da condição corporal variando de 3,0 a 3,5, em escala de 1 a 5, onde 1 corresponde ao animal muito magro e 5 ao muito gordo (Dirksen et al., 1993). As novilhas foram mantidas em sistema de pastejo rotacionado, com gramínea de Brachiaria brizantha cv. Marandu, e suplementadas com mistura mineralizada. O desenvolvimento folicular foi acompanhado através de exames ultra-sonográficos, efetuados a cada 24 horas. Foi utilizado aparelho de ultra-som equipado com transdutor linear endoretal bifreqüencial 5.0-7.5 MHz. No momento do exame, o transdutor foi movimentado sobre a superfície dos ovários e, quando apropriado, a imagem do monitor foi congelada para a mensuração dos folículos e do corpo lúteo, se presente. Durante cada exame foram desenhados diagramas com o posicionamento relativo dos folículos de diâmetro igual ou maior a quatro mm (³4 mm). Quando a imagem do folículo acompanhado não apareceu esférica, o diâmetro foi estimado pela média do maior e menor diâmetro (Savio et al., 1988), sendo o dia zero considerado o dia da detecção de um ou mais folículos ³4 mm. As variáveis analisadas foram: Comprimento de onda (dias) - equivale ao número de dias entre o dia da detecção do folículo (dominante ou maior subordinado) até o último dia de sua detecção, última mensuração efetiva (³4 mm); Tamanho máximo (mm) - folículo (dominante ou subordinado); Crescimento (dias) ou fase de crescimento - equivale ao número de dias entre a detecção do folículo (dominante ou subordinado) ³4 mm, ou corpo lúteo, e o dia que o mesmo cessa seu crescimento progressivo (apresentando um diâmetro igual ao anteriormente medido); Taxa de crescimento (mm/dia) - o tamanho mínimo do folículo (³4 mm), efetivamente mensurado, subtraído do tamanho máximo, dividido pelo número de dias de crescimento; Platô (dias) ou fase estática - se estende do último dia da fase de crescimento (quando começa a apresentar diâmetros repetidos) até quando o folículo (dominante ou subordinado) ou corpo lúteo, inicia uma diminuição contínua no seu diâmetro (apresentando um diâmetro diferente, menor que o anteriormente medido); Dia inicial da atresia ou luteólise - dia que o folículo (dominante ou subordinado) ou corpo lúteo começa a diminuição progressiva em seu crescimento (diâmetro é menor que o anteriormente medido); Atresia (dias), luteólise ou fase de regressão - equivale ao número de dias entre o dia inicial da atresia e o último dia de detecção do folículo (dominante ou subordinado), ³4 mm, ou corpo lúteo; Taxa de atresia (mm/dia) ou luteólise - o tamanho máximo do folículo (dominante ou subordinado) ou corpo lúteo, subtraindo o tamanho mínimo efetivamente mensurado (³4 mm) no final da fase de atresia ou regressão, dividido pelo número de dias desta fase. Os dados foram submetidos à análise de variância pelo procedimento GLM (SAS, 1995). A significância das diferenças entre médias foi determinada pelo teste t de Student, com nível de probabilidade de 5%. Resultados e DiscussãoO desenvolvimento folicular, caracterizado pelo crescimento e regressão do folículo dominante, manifestou-se sempre de forma semelhante a ondas, cada uma delas apresentando um folículo dominante e um número variável de outros menores (subordinados). Esta característica é semelhante ao que foi relatado por Adams et al., (1994) e Evans et al., (1994), ao estudarem novilhas Hereford pré-púberes; por Figueiredo et al., (1997) em novilhas Nelore; Gambini et al., (1998) estudando vacas da raça Gir e por Borges et al., (2001) que utilizaram novilhas mestiças Holandês x Gir. Durante o período de estudo não foram observados ovulação e corpo lúteo nas fêmeas estudadas, entretanto todas elas apresentaram padrão semelhante de desenvolvimento folicular. Foi observada a ocorrência de sucessivas ondas, que apresentaram-se altamente variáveis em comprimento (duração em dias). Por esta razão foram categorizadas em comprimento de ondas um (CO1) £12 dias; dois (CO2), entre 13 e 16 dias e três (CO3) ³17 dias, para serem melhor analisadas. Foram acompanhadas 49 ondas, sendo seis CO1 (12,2%), 31 CO2 (63,3%) e 12 CO3 (24,5%). As características dos folículos dominantes e maiores subordinados, para CO1, CO2 e CO3, podem ser observadas na Quadro 1 e na Figura 1. Nas ondas com CO1 o diâmetro máximo do folículo dominante foi significativamente menor (9,4±0,3 mm) que CO2 (10,4±0,2 mm) e CO3 (11,6±0,4 mm), (P<0,05). Estes valores são similares àqueles das ondas de crescimento folicular anovulatórias, para intervalos interovulatórios, com duas e três ondas, informados em estudos com raças zebuínas por Figueiredo et al., (1997) que encontraram 11,3±0,4 mm para o primeiro folículo dominante em fêmeas de duas ondas e 10,4±0,3 e 9,4±0,3 mm para o primeiro e segundo folículo dominante, respectivamente, em animais de três ondas foliculares. Gambini et al. (1998), trabalhando com vacas da raça Gir, encontraram valores de 11,2±0,7 mm para o primeiro folículo dominante em vacas de duas ondas e 12,2±0,8 e 10,8±0,9 mm para o primeiro e segundo folículos dominantes, respectivamente, em vacas com três ondas foliculares. Todos os valores relatados por estes autores referem-se a folículos dominantes de ondas anovulatórias. Por sua vez Borges et al. (2001) estudando a dinâmica folicular em novilhas cíclicas mestiças Holandês/Zebu, relataram que o diâmetro máximo do primeiro FD, em animais de duas ondas, foi igual a 7,1±1,2 mm e para animais com três ondas os valores foram de 6,8±1,4 e 6,2±0,9 mm, para o primeiro e segundo folículo dominante, respectivamente, valores inferiores aos encontrados neste estudo. Já Sávio et al. (1988) encontraram 14,25 mm para o primeiro folículo dominante em animais de duas ondas e 15,9 mm para o segundo em fêmeas taurinas de três ondas, enquanto Sirois & Fortune (1988) observaram 12,0 mm como tamanho máximo do primeiro folículo dominante, em vacas Holandesas de duas ondas e 12,3 e 10,22 mm para o primeiro e segundo folículos dominantes, respectivamente, naquelas com três ondas. Quadro 1. Comparação das características (média±EPM) dos folículos dominante e maior subordinado, observadas em novilhas Gir, com diferentes comprimentos de ondas anovulatórias.
a,b,cMédias seguidas de mesma letra em cada linha não diferem (P>0.05), para folículos dominante e maior subordinado. Das 49 ondas estudadas, seis (12,2%) apresentaram CO1, 31 (63,3%) CO2 e 12 (24,5%) tiveram comprimento ³17 dias (CO3). Durante o período do estudo 10 (76,92%) novilhas apresentaram quatro ondas, com diferentes comprimentos, enquanto três (23,08%) manifestaram somente três ondas de crescimento folicular. O valor encontrado neste estudo, para o diâmetro máximo do folículo dominante (11,6±0,4 mm), foi ligeiramente menor que os 12,0±0,3 mm relatados por Adams et al. (1994), em 11 animais Hereford prépúberes. Entretanto, o comprimento médio desta onda (15-16 dias) pode ser enquadrado melhor em CO2. Evans et al. (1994), trabalhando também com novilhas da raça Hereford, encontraram valores de 13,7±0,5 mm como tamanho máximo do folículo dominante da última onda anovulatória anterior à primeira ovulação. Para as demais características do folículo dominante, como o platô e as taxas de crescimento e atresia, entre os três grupos de CO não mostraram-se diferenças significativas (P>0,05). Para os folículos subordinados, as propriedades estudadas não diferiram significativamente (P>0,05) entre as três classes de CO. Os resultados, referentes à fase de crescimento e à taxa de atresia dos folículos dominante e maior subordinado, para CO1 e CO2, obtidos neste estudo, são similares aos valores relatados por Adams et al. (1994). Entretanto, a taxa de crescimento e dias até o platô encontrados foram inferiores aos informados por estes autores. Na Figura 1 estão representados os números médios de folículos pequenos (d»5 mm), médios (³6 e <8 mm), grandes (³8 mm) e total de folículos, para CO1, CO2 e CO3. No início do período de observação, entre os dias 2 e 3, para todos os COs, houve uma diminuição no número de folículos pequenos e um aumento no número de folículos médios. Por volta dos dias 1-2 (CO1), 2-3 (CO2 e CO3), do período de observação, notou-se um pequeno aumento do número de folículos grandes, acompanhado da diminuição dos médios, o que representa momento de dominância folicular. O número de folículos pequenos manteve-se diminuído até por volta dos dias 5 a 6 (CO1) e 7 a 8 (CO2 e CO3), quando voltou a aumentar, caracterizando recrutamento de novos folículos. A curva de folículos totais tendeu a comportar-se de maneira semelhante àquela dos folículos pequenos, uma vez que estes são em número significativamente maior que os médios e grandes, apresentando semelhança àquela encontrada no trabalho de Adams et al. (1994). As tendências das curvas sobre o número de folículos pequenos, médios e grandes comportaramse igual às curvas descritas no trabalho do Gambini et al. (1998), durante o período de dominância folicular. Neste aspecto, pode-se observar que o folículo dominante diminuiu o ritmo de crescimento dos folículos subordinados e impediu o recrutamento de um novo grupo de folículos, como reportado por Ko et al. (1991). De acordo com Turzillo e Fortune (1993) o folículo dominante secreta inibina, que causa retroalimentação negativa diminuindo a secreção de FSH, essencial para o recrutamento e o desenvolvimento folicular. No entanto, segundo Ginther et al. (1996) o folículo dominante, mudando a ênfase na dependência de gonadotrofina, continua a crescer e a se desenvolver porque torna-se LHdependente, enquanto os outros folículos subordinados são privados de FSH. Considerando que os animais apresentavam boa condição corporal, entre 3,0 e 3,5, isto permite supor que este padrão de crescimento folicular seja o normal para esta raça, na idade apresentada pelos animais durante o período de estudo (22±2 meses). Apesar da variação de quatro meses na idade das novilhas, os autores acreditam que esta pequena diferença não interferiu nos resultados encontrados, pois o padrão de crescimento folicular foi uniforme entre os animais estudados. ConclusõesO desenvolvimento folicular de novilhas prépuúberes da raça Gir caracterizou-se por sucessivas ondas de crescimento folicular, observando-se diferentes comprimentos de onda em um mesmo animal, mostrando-se similar aos resultados de estudos em novilhas pré-púberes de outras raças zebuínas. A dinâmica folicular caracterizou-se predominantemente por duas e três ondas de crescimento folicular, havendo variação no número de ondas de um intervalo interovulatório para outro. O padrão da dinâmica folicular, no período imediatamente anterior à puberdade, na raça Gir permite concluir que este é similar ao encontrado em outras raças zebuínas e taurinas, constituindo informação importante para o estudo da foliculogênese após o estabelecimento da ciclicidade estral. Literatura Citada
© 2005 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la05007f1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}