 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 13, No. 2, Mayo-Agosto, 2005, pp. 56-62 Efecto del recuento de células somáticas sobre la longevidad funcional del ganado frisón analizado mediante técnicas de análisis de supervivencia. Effect of somatic cell scores on functional longevity of dairy cattleassessed by survival analysis. Z. Chirinos1, D. Hernández2, y M.J. Carabaño3. Facultad de Agronomía, Universidad del Zulia. Apartado 15205, Maracaibo 4005-A. Recibido Enero 7, 2005. Code Number: la05008 ABSTRACT: The objective of this study was to analyze the effect of somatic cell scores on functional longevity (FL) assessed using survival analysis. Longevity was measured as days of productive life. Functional longevity was obtained by global longevity correcting for milk production. Data of production and somatic cell count (SCC) consisting of records from 20.359 Spanish Holstein cattle, with date of first calving between 1988 and 2000. Factors included in the Weibull model were: herd-year-season of calving (HYS), age at first calving (AFC), lactation number-stage of lactation (NS), annual change in herd size (CHZ), milk (MY), fat (FY) and protein (PY) yield. All the effects were significant (P<0.0001) with HYS and NS those as that more contributed to explain the differences on the relative risk of culling (RR), followed by MY and contributed to explain the differences on the relative risk of culling (RR). An increase in the SCC levels caused a greater RR. An important effect of SCC and udder depth in FL was observed. Cows with greater SCC had almost 4.0 times more RR and it was more evident in very deep udders animals. Key words: Longevity, somatic cell count, survival analysis. RESUMEN: El presente trabajo tiene como finalidad estudiar el efecto del recuento de células somáticas (RCS) sobre la longevidad funcional (LF), analizado mediante técnicas de análisis de supervivencia. La longevidad funcional se midió como días de vida productiva, obtenida mediante la corrección de la longevidad global por producción lechera. Se utilizaron datos de longitud de vida productiva, producción lechera y recuento celular de 20.359 de ganado Holstein español que hubiesen tenido su primer parto entre 1.988 y 2.000. Se utilizó un modelo de Weibull que incluía los efectos de rebaño-año-época de parto (RAE), edad al primer parto (EP), número de lactación-estado de la lactación (NEL), cambio anual en el tamaño del rebaño (CTR), producción de leche (PL), grasa (PG), proteína (PP), y RCS. Todos los efectos resultaron significativos (P<0,0001), siendo RAE y NEL los que más contribuyeron a explicar las diferencias sobre el riesgo relativo de desecho (RR), seguidas por PL. Un incremento en los niveles de RCS provocó un mayor RR. Se observó un efecto importante de RCS y profundidad de ubre en LF, las vacas con mayor RCS presentaron casi 4,0 veces más RR y fue más evidente en animales con ubres muy profundas. Palabras clave: Longevidad, recuento de células somáticas, análisis de supervivencia. Introducción La longevidad es un carácter económicamente importante en ganado vacuno lechero, tanto por el interés que tiene per se el aumentar la vida productiva de los animales y con ello disminuir el costo de la reposición, como por su relación con la capacidad de los animales de evitar el desecho involuntario. El desecho involuntario es aquel que el ganadero debe hacer sin que le reporte ningún beneficio, a diferencia del desecho voluntario por baja producción lechera que implica una mejora del nivel de producción de la explotación (Ducrocq et al., 1988). Una de las primeras causas de desecho involuntario en las explotaciones lecheras es la susceptibilidad de los animales a desarrollar infecciones mamarias. La importancia económica de este tipo de infecciones es elevada por el aumento de gastos veterinarios, la bajada de producción lechera y la necesidad de descartar leche no apta para el consumo humano, los posibles daños permanentes en la ubre, etc. Existen varias medidas en los sistemas rutinarios de control lechero que están relacionadas con la susceptibilidad al desarrollo de infecciones mamarias. Las medidas más comunes son el recuento de células somáticas en leche (RCS), medidas relacionadas con la morfología de la ubre, y, datos de incidencia de mastitis clínica. En cuanto a la morfología de la ubre, el carácter más claramente asociado a las infecciones mamarias o a un RCS elevado es la profundidad de ubre (Charffeddine et al., 1997, Rupp y Boichard, 1999). En la población Frisona española, tanto el dato de RCS como los caracteres de calificación morfológica se recogen de forma habitual en los animales en control lechero, mientras que el dato de mastitis clínica no suele estar disponible. La relación entre el desarrollo de infecciones mamarias y la longevidad ha sido investigada previamente en varios estudios, tanto en la población Frisona española (Charffeddine, 1998) como en otras poblaciones de ganado vacuno lechero (Rogers, 2002; Samoré, 2003). En la mayoría de los casos, esta relación ha sido cuantificada a través de la estima de la correlación entre la longitud de la vida productiva y el RCS, obtenida mediante el uso de modelos lineales bi-carácter. En general, las estimas obtenidas muestran valores negativos, indicando un antagonismo entre la longevidad y el RCS (Charffeddine, 1998; Roxström y Strandberg, 2002). Es decir, la predisposición a mostrar un RCS alto está relacionada con una longevidad más baja. Los valores de las estimas obtenidas son, sin embargo, relativamente bajos, entre –0,30 (Rogers et al., 1998) y –0,32 (Mrode et al, 2000). Estimas de similar magnitud han sido encontradas entre el carácter profundidad de ubre y diferentes medidas de longevidad (Vollema, 1998). Dado que la correlación es una medida de la relación lineal entre dos variables, la existencia de relaciones no lineales entre las variables podría explicar, al menos en parte, estos bajos valores. Por otra parte, los datos utilizados en esos estudios correspondían a animales con datos completos de longevidad, excluyéndose datos de animales aun no desechados. Esto supone una selección no aleatoria de los datos utilizados en el análisis, que puede provocar sesgos en las estimas de los parámetros genéticos. Por último, el uso de determinados modelos no lineales como los empleados con las técnicas de análisis de supervivencia (e.j. Kalbfleisch y Prentice, 1980) se considera como teóricamente más adecuado para el análisis de datos de longevidad (e.j. Ducrocq et al., 1988). Estas técnicas han sido empleadas para estudiar el impacto que el recuento de células somáticas y la mastitis tienen sobre la longevidad funcional en otras poblaciones de ganado vacuno lechero (Beaudeau et al., 1995; Samoré, 2003; Caraviello et al., 2005). Separadamente, la relación entre la longevidad funcional y los caracteres de morfología mamaria, y, en particular, la profundidad de ubre ha sido también investigada con técnicas de análisis de supervivencia en varias poblaciones de ganado vacuno lechero (Büenger et al., 2001; Larroque y Ducrocq, 2001; Chirinos, 2003) El presente trabajo tiene como finalidad estudiar conjuntamente el efecto del recuento de células somáticas y de la profundidad de ubre sobre el riesgo de desecho de la población Frisona española analizada mediante las técnicas de análisis de supervivencia. Materiales y Métodos Datos Los datos de longevidad, Recuento de células somáticas (RCS) y producción lechera de los animales que participan en este estudio proceden del control lechero oficial de la Comunidad Autónoma Vasca, España. La longevidad se definió como longitud de vida productiva (LVP) para cada animal y fue calculada como el número de días entre la fecha del primer parto y la fecha del último pesaje de leche registrado. Esta fecha fue usada como fecha de eliminación o censura, dado que no se dispone de datos fiables de causa y fecha de baja de las vacas en los rebaños. Se consideró que el dato de longevidad era completo (es decir de un animal que realmente había sido desechado) si existía una diferencia superior a 7 meses desde la fecha de su último pesaje de leche y la fecha del último pesaje de leche del rebaño. En caso contrario, el dato de LVP era considerado como censurado o incompleto, es decir de un animal que aun no había sido desechado en la fecha del último pesaje controlado. Asimismo, cuando el animal era llevado a otro rebaño, sólo se contabilizó en el cálculo de la vida productiva la primera parte de la información registrada fue incluida en el estudio, descartándose la información procedente de la segunda parte en otro rebaño y el dato se consideró como censurado. Siguiendo los patrones establecidos por la Confederación de Asociaciones de Frisona española (CONAFE) para sus evaluaciones genéticas, no se incluyeron en el estudio datos de todas aquellas vacas con fecha de nacimiento faltante, sin información de primer parto, con edad al primer parto fuera del intervalo 540 – 1.220 días, con un intervalo entre partos menor a 305 días, y sin datos de producción o producciones fuera de los rangos 3.000 – 20.000 Kg. para producción de leche, 1,5% - 9,0% para porcentaje de grasa y 1,0%-7,0% para porcentaje de proteína. Animales con fecha de primer parto entre 1.988 y 2.000. Se consideraron únicamente rebaños con al menos 3 años de información y con al menos 20 vacas controladas a lo largo de los años de estudio. Asimismo, para garantizar una precisión mínima en la estima del efecto grupo de comparación, se descartaron datos de clases de rebaño-año con menos de 5 registros y se exigió un mínimo de dos datos completos en las clases año-época de parto y en las clases de rebaño. Los datos fenotípicos de RCS corresponden a la media de las puntuaciones para el recuento de células somáticas transformadas de todos los pesajes, exigiendo como mínimo 8 pesajes y 310 días de duración de lactación. Las puntuaciones de RCS en cada pesaje se corrigieron con las soluciones BLUP por los efectos del mes de pesaje y el estado de lactación obtenidos del sistema de evaluación genética de RCS en la Comunidad Autónoma Vasca. Análisis estadístico El análisis fue realizado con el paquete estadístico Survival Kit de Ducrocq y Sölkner V3.12 (Ducrocq y Sölkner, 1998; Sölkner y Ducrocq, 1999), utilizando el siguiente modelo de referencia: h(t)=ho{RAE i ( τ )+ EP j + NEL k ( t , t ’)+ CTR l (τ)+PLm(τ)+PGn (τ)+PPo(τ)+IRC p +RCSq} donde, h(t)= función de riesgo (probabilidad de un animal de ser eliminado t días después del primer parto), ho(t)= λρ(λt)ρ-1= función de riesgo base de una vaca, bajo la distribución de Weibull con los parámetros de forma, ρ, y escala, λ. RAEi(t)=Efecto del rebaño-año-época de parto, con cambios en el primer día de los meses de enero, abril, julio y octubre de cada año, EPj = Efecto de la edad al primer parto en meses. Se consideraron 9 clases: 1 (≤ 24,0), 2 (24,1; 25,0), 3 (25,1; 26,0), 4 (26,1; 27,0), 5 (27,1; 28,0), 6 (28,1; 29,0), 7 (29,1; 31,0), 8 (31,1; 34,0), 9 (>34,0). NELk(t,t’)= Efecto del número de lactación (1,2,3,4,5,6, ≥7), que cambia en cada fecha de parto* estado de la lactación, con cambios a 0, 30, 60, 90, 180, 240 y 300 días de cada lactación, CTRl(τ)= Efecto del cambio anual en porcentaje del tamaño del rebaño, con cambios en el primer día del mes de enero de cada año. Se consideraron 11 clases: 0 (desconocida), 1 (≤ - 30%), 2 (-30%, -20%), 3 (20%, -10%), 4 (-10%, 0%), 5 (0%), 6 (> 0%, 10%), 7 (10%, 20%), 8 (20%, 30%), 9 (30%, 40%), 10 (> 40%). La clase 0 fue asignada tomando en cuenta los siguientes aspectos: combinaciones de rebaño-año con menos de 11 datos, el primer año y los dos últimos años que el rebaño aparecía en el registro lechero, y rebaños que incrementaban en más de un 200% el número de animales de un año a otro. Este grupo de datos asignados a la clase cero no entraron en el cálculo del porcentaje de variación anual. PLm(τ), PGn(τ) y PPo(τ)= Efecto de la producción de leche, grasa y proteína con cambios al inicio de cada lactación. Los datos de producción vienen ajustados o corregidos con las soluciones BLUP, de los efectos número de lactación (1er parto), edad al parto (24 meses) y mes de parto (Enero) del sistema de evaluación nacional de los caracteres de producción. Las clases para estos efectos se asignaron en base a la desviación porcentual de la producción del animal con respecto a la media de producciones de animales en rebaño-año de parto. Se consideraron 8 clases: 1(< -30%), 2 (-30%, -20%), 3 (-20%, -10%), 4 (-10%, 0%), 5 (0%, 10%), 6 (10%, 20%), 7 (20%, 30%), 8 (> 30%). IRCp = Efecto presencia o ausencia de información de recuento. RCS = Dato de recuento de células somáticas q (x1000/ml), con cambios en cada lactación. Se agruparon en 5 clases: 0 (desconocida, para animales sin datos de recuento), 1 (< 100), 2 (≥100;<200), 3 (≥200; <400), 4 (≥400; <800), 5 (≥ 800). Los caracteres de producción de leche fueron incluidos en el modelo que se denominó LF, de longevidad funcional y excluidos en el modelo denominado LG, de longevidad global. En un segundo modelo, se incluyó también el efecto de la profundidad de ubre en forma individual y como interacción con RCS para estudiar la posible asociación entre ésta y la longevidad funcional. La variable profundidad de ubre viene medida en nueve categorías que van desde ubres muy profundas (clase 1-3), deseables (4-6) a ubres muy poco profundas (clase 7-9). Para cuantificar la magnitud de la contribución de cada factor sobre la longevidad funcional se recurrió al test de razón de verosimilitud, que permite estudiar la significación global del modelo y de cada una de las variables que se incluyen en él y se efectúa a partir del incremento de la verosimilitud al incorporar k variables en el modelo. El estadístico utilizado -2logL sigue la ley de χ2 con k grados de libertad proporcionado por el programa Survival Kit. El estadístico R2 de Maddala, similar al tradicional R2 de los modelos de regresión lineal, fue también otro criterio considerado para evaluar el aporte de los distintos factores y la bondad de ajuste global del modelo. En cuanto a la comparación de los diferentes modelos propuestos para evaluar la inclusión de la información de RCS y profundidad de ubre sobre la longevidad funcional, se utilizaron también los criterios de Akaike’s (Akaike,s Information Criterion, AIC; Akaike, 1974) y el BIC (Bayesian Information Criterion; Schwarz, 1978). El modelo más adecuado es el aquel que minimice AIC y el BIC. AIC= -2logL+2k BIC= -2logL+klog(n) donde, k= número de soluciones estimables del modelo y n= número de datos completos. Por otra parte, para evaluar el tipo de relación entre el RCS, la profundidad de ubre y el riesgo de desecho se obtuvieron las estimas máximo verosímiles proporcionadas por el programa bajo los modelos LG y LF. Resultados y Discusión
En el Cuadro 1 se presenta una descripción de las características más importantes de los datos utilizados en este estudio, con detalles de la cantidad, media, moda, mediana, desviación estándar, mínimo y máximo de longevidad, recuento de células somáticas y profundidad de ubre. Del total de 20.359 datos de longevidad, 7.949 (39.04%) fueron censurados o incompletos. Nuestros datos son comparables a los reportados por Caraviello et al.,(2005), y están por debajo a los reportados por Samoré (2003) quien obtuvo un 57%. Según Vukasinovic et al., (1999) el número mínimo de registros censurados que son aceptables para obtener resultados confiables debería estar entre los 30 y 40%. En cuanto a la significación de los efectos incluidos en el modelo, en el Cuadro 2 se exponen los resultados del test de razón de verosimilitud donde se compara el modelo total contra modelos que van excluyendo un efecto cada vez. Todos los efectos ambientales diferentes al recuento de células somáticas incluidos en el modelo mostraron un efecto significativo (P <0,0001) sobre la longevidad funcional de acuerdo con el test χ2. Los efectos que mostraron una contribución más alta a la verosimilitud (mayor χ2) fueron RAE, NEL, siguiendo en importancia PL. Es decir, junto con las condiciones de manejo asociadas a cada rebaño en un periodo de tiempo determinado, estos son los principales factores que determinan el riesgo relativo de un animal de ser desechado. En contraste con el desecho por producción de leche, la producción de grasa y proteína, aunque con un efecto significativo, tienen un menor impacto frente a la producción total de leche en el desecho de los animales. Estos resultados concuerdan con otras investigaciones en las que se estudiaban el efecto de factores sistemáticos en la longevidad funcional (Vukasinovic et al., 1997; Durr et al.,1999). El estadístico R2 de Maddala para el modelo completo, similar al tradicional R2 de los modelos de regresión lineal, muestra que la bondad del ajuste de este modelo es elevada 0,66, considerándose que los factores considerados identifican en gran parte las condiciones ambientales que determinan el riesgo de los animales de ser desechados en la población analizada. Las estimaciones de los parámetros de Weibull (ρ e intercepto) para los diferentes modelos de LF propuesto, se presentan en el Cuadro 3. Observamos valores de ρ entre 2,32 y 2,41. Un valor estimado de ρ superior a 1, indica que el riesgo base incrementa con el tiempo, lo que parecería lógico para el ganado vacuno de leche si consideramos que el riesgo de desecho incrementa con la edad del animal porque se vuelven más susceptibles a los efectos degenerativos propios del proceso de envejecimiento (Boettcher et al., 1999). En evaluaciones hechas en Québec Durr et al., (1999) reportaron un ρ=1,62 y en Suiza Vukasinovic et al., (1999) estimaron un ρ=1,53 muy por debajo a los encontrados en este estudio. El intercepto de la distribución de Weibull (ρ logλ), es una medida de la varianza asociada al efecto de la combinación rebaño-año-época de parto y nos indica la homogeneidad o heterogeneidad en las prácticas de desecho entre los ganaderos (Chirinos, 2003). Con nuestros datos las estimaciones de este parámetro presentaron un rango de –15,33 y –15,79. Esto nos señala que existe poca diferencia entre los criterios que siguen los ganaderos en la forma de manejar el rebaño y a la posible influencia de los efectos ambientales. A pesar de ello, esta tendencia está por encima a lo reportado en rebaños de frisón en Québec (Durr et al.,(1999) e Italia (Samoré, 2003) con valores para el intercepto de -11,18 y -11,20 respectivamente. Cuadro 1. Descripción de los datos utilizados en este estudio.
RCS=Recuento de células somáticas; D.S= Desviación estándar Cuadro 2. Resultados del test de razón de verosimilitud comparando el modelo completo con modelos que excluyen un efecto cada vez.
RAE=Rebaño*Año*Epoca; EP=Edad al 1er parto; IRC= Presencia o ausencia de dato de recuento; RCS= Recuento de células somáticas; NEL=Lactación*Estado de la lactación; CTR= Cambio en el tamaño del rebaño; PL=Producción de leche, PG=Producción de grasa; PP=Producción de proteína. Cuadro 3. Parámetros de Weibull estimados (r e intercepto) asociados a los modelos alternativos de longevidad funcional (LF).
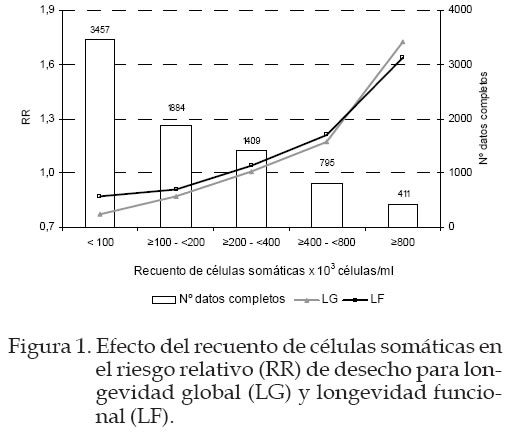
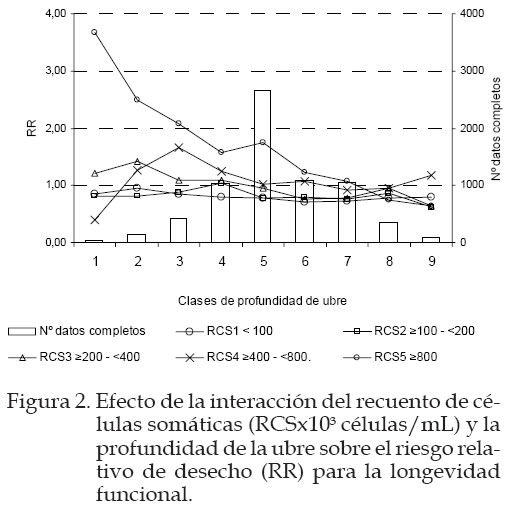
La inclusión de la información de RCS y profundidad de ubre representó mejoras significativas en el ajuste del modelo para explicar las causas de desecho en el ganado Frisón, tal como se muestra en el Cuadro 4, cuando comparamos el modelo básico con los otros propuestos. En el Cuadro 4 se presenta el valor de –2 log L, el R2 de Maddala (semejante al R2 de los modelos lineales), y los criterios de Akaike (AIC) y el criterio bayesiano (BIC) para el modelo LF. Resultados similares se obtuvieron sobre LG. En ambos casos el mejor modelo que explica la influencia del recuento sobre la longevidad funcional fue el tercer modelo que consideraba la interacción de PU*RCS con valores de AIC=196.720 y para el BIC= 251.629 respectivamente. En las Figuras 1 y 2 se presenta la estima del efecto de cada clase del carácter de RCS y RCS*PU, respectivamente, sobre el riesgo relativo de desecho (RR), con relación a una clase de referencia que presenta un riesgo relativo que vale 1 y corresponde a la clase con mayor número de datos completos contra las que se comparan todas las clases, de acuerdo a los principios del Survival Kit. En este caso, la clase 3, de nivel intermedio de RCS para LG y LF y la clase 5 para la interacción RCS*PU. En las figuras se ha incluido también el número de datos completos en todas las clases como indicador de la fiabilidad de las estimas encontradas. Cuadro 4. Contribución a la verosimilitud (-2logL) de los factores de recuento de células somáticas (RCS) y de su interacción con la profundidad de ubre (PU), el estadístico R2 de Maddala, los criterios de Akaike (AIC) y el bayesiano (BIC) asociado a los modelos alternativos de longevidad funcional (LF).
AIC= -2logL+2k; BIC= -2logL+klog(n) donde, k= número de soluciones estimables del modelo y n= número de datos completos. Según puede verse en la Figura 1, no existieron diferencias en las tendencias de RR para LG y LF, observándose un aumento en el RR de desecho a medida que aumenta el nivel de RCS. Esto indica que el nivel de RCS influye en la longevidad básicamente a través de una disminución de la funcionalidad de los animales. El RR frente a la clase de referencia (e»200; <400 x 103 célulasl/mL) osciló entre 0,77 y 1,72 para LG y entre 0,87 y 1,63 para LF. Así, los animales con un nivel de RCS inferior a 100x103 células/mL tienen un riesgo de desecho 2,23/1,87 veces inferior al riesgo de desecho de los animales con el nivel máximo de RCS, superior a 800 x 103 células/ mL para el caso de LF/LG. Estos resultados coinciden con los de Beaudeau et al., (1995). Ellos reportaron que vacas con una concentración de RCS superior a 800.000 células/ml tenían 1,7 veces más riesgo de ser eliminadas que una vaca que tuviese un valor por debajo de 300.000 células/mil. Samoré (2003), aunque utilizó una escala diferente a la nuestra, encontró en su investigación que las vacas ubicadas en las clases más altas para RCS tenían casi 3 veces mas riesgo de desecho que las que ocupaban la clase de referencia, mientras que los animales en clase de mínimo RCS tenían la mitad de riesgo que los animales de referencia. Según Caraviello et al., (2005) vacas con lactaciones que mantenían un mayor RCS (>700.000 células/mL) comparada con vacas de un RCS promedio (200.000-250.000 células/mL) poseían un RR de 2 a 4 veces mayor y de 10 a 25 veces por encima si el desecho era debido a mastitis. Otro aspecto interesante reflejado en la Figura 1 es la relación no lineal entre el RCS y el RR de desecho. En esta Figura puede observarse un punto de cambio de pendiente para los animales con un nivel máximo de RCS. Este punto coincidiría con el umbral de RCS a partir del cual se considera que el animal padece una infección clínica (Schukken, et al., 1997). En la Figura 2 se muestra el efecto de las nueve clases de profundidad de ubre sobre el RR para cada uno de los cinco niveles de RCS. Según puede observarse, el riesgo de desecho no varía de forma notable con la profundidad de ubre excepto para animales con un RCS alto (≥800.000 células/mL). Así, animales con RCS elevado y ubres muy profundas tienen casi 4 veces más riesgo que los animales con RCS elevado y ubres poco profundas. Los animales con ubres profundas y recuentos altos son los que van a estar en riesgo alto mientras que animales con ubres poco profundas tienen menos riesgo. Roger et al., (1998) determinaron que existía una correlación genética moderada y positiva entre profundidad de ubre y RCS para vacas de Dinamarca y Suecia de 0,37 y 0,52 respectivamente, e indicó que esta medida sería muy útil a la hora de considerar la sanidad de la ubre, especialmente cuando los datos de mastitis no procedían de vacas registradas. Las vacas con ubres más profundas tienen una mayor predisposición a contraer infecciones mamarias por la mayor facilidad de contacto con los agentes infecciosos y por la mayor propensión a lesiones de la ubre. Si, aparte de tener una morfología que facilita la entrada de agentes infecciosos en la ubre, se trata de animales con menos capacidad de defensa frente estos agentes, el riesgo de contraer enfermedades mamarias y, por tanto, de ser desechadas por este motivo se multiplica. Conclusiones El nivel de RCS en leche tuvo un efecto importante sobre el desecho de animales, en el sentido de que animales con el nivel más alto de RCS tienen un riesgo de desecho cercano al doble del riesgo que presentan animales con los niveles de RCS más bajo. Esta relación no es lineal, siendo más notable el efecto para RCS superiores a 800.000 células/mL. Además, el riesgo de desecho se multiplica casi por cuatro cuando se trata de animales con RCS alto y ubres más profundas frente a animales con ubres poco profundas y niveles bajos o intermedios de RCS. Literatura Citada
© 2005 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la05008f2.jpg] [la05008f1.jpg] |
| |||||||||

{kind=link}
{kind=link}