 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Archivos Latinoamericanos de Produccion Animal, Vol. 14, No. 2, 2006, pp. 33-41 Aceitação de larvas de diferentes grupos genéticos de Apis mellifera na produção de abelhas rainhas Acceptance of larvae from different genetic groups of Apis mellifera in queen bee production V.N. Albarracín1, S.R. Cunha Funari, E.M. Romero Arauco, R. de Oliveira Orsi Departamento de Produção e Exploração
Animal, Faculdade de Medicina Veterinária e Zootecnia, Recibido Marzo 29, 2005. Aceptado Enero 13, 2006. Code Number: la06005 ABSTRACT. Honeybee renewal depends on the reproductive process, in which the queen plays a dominant role, thus fostering beehive survival and maintenance of hive characteristics and productivity. Royal jelly, secreted by the hypopharyngeal and mandibular glands of worker honeybees, constitutes the queen's diet, both in the larval stage and the adult. The goal of this study was to evaluate the percentage acceptance of larvae, of different genetic groups of Apis mellifera, in the production of queens. The work was carried out at the Beekeeping Section, FMVZ, UNESP, Campus de Botucatu Sp Brasil. Three treatments were included: T1 (larvae from the same genetic group and the same colony), T2 (same genetic group and different colony), and T3 (different genetic groups). The two genetic groups used were Africanized and Italian. Queen cells were placed in three different bar positions: A (in front, close to the entrance), M (in the middle), and F (at the rear, far from the entrance). In the process of queen production, the genetic groups and treatments showed no significant differences in percentage acceptance of larvae and weight of royal jelly for queen cells. Bar positions had no significant effect on the acceptance percentage of larvae, but the M position resulted in greater weight of royal jelly for queen cells than positions A and F. Greater percentage of larvae acceptance was associated with greater weight of royal jelly for queen cells and greater weight of larvae produced, in both genetic groups. Key word: Apis mellifera, queen production, larvae RESUMO. A colônia de abelhas Apis mellifera se perpetua por meio de sua contínua renovação, onde a rainha desempenha um papel de suma importância na sobrevivência da colônia e continuidade da espécie, determinando as características e a produtividade dessa colmeia. A rainha é alimentada, durante as fases larval e adulta, com geleia real secretada pelas glândulas hipofaringeanas e mandibulares das operárias nutrizes. O objetivo deste trabalho foi avaliar a porcentagem de aceitação de larvas nos diferentes grupos genéticos de abelhas Apis mellifera (italiana e africanizada), na produção de rainhas. O experimento foi desenvolvido no Setor de Apicultura, Facultade de Medicina Veterinaria e Zootecnia, UNESP, Campus de Botucatu, SP, Brasil. Foram estudados 7 colmeias do grupo genético africando e 7 colmeias do grupo genético italiano, distrubuidas em três tratamentos: T1 (larvas do mesmo grupo genético e matrilínea,). T2 (larvas do mesmo grupo genético e diferente matrílinea) e T3 (larvas de diferente grupo genético), sendo que as realeiras estavam posicionadas em três locais da barra do quadro porta realeiras: A (anterior, na parte mais próxima do alvado), M (centro) e F(na parte mais distante do alvado). Conclui-se que na produção de rainhas, os grupos genéticos e os tratamentos não apresentaram diferenças significativas na porcentagem de aceitação de larvas e na produção da geleia real por realeira; quanto aos locais, não houve diferença significativa para a porcentagem de aceitação de larvas, mas quanto ao peso da geleia real, o local M apresentou melhor produção que os locais A e F; e quanto maior foi a porcentagem de aceitação de larvas, maior foi o peso da geleia real e maior o peso da larva, para ambos grupos genéticos. Palavras chave: Apis mellifera, rainhas, produção de rainhas, larvas Introdução A colônia de abelhas se perpetua por meio de sua renovação, sendo a rainha responsável pela sobrevivência e continuidade da colônia e da espécie (Silva, 2000), influenciando nas características e na produtividade das colmeias. A rainha de Apis mellifera sofre cinco ecdises durante seu desenvolvimento, de ovo a adulta, sendo 3 dias como ovo; 5.5 a 6 dias como larva; 6.5 a 7 dias com pré -pupa e pupa. Na fase, de larva a imago, os fatores meteorológicos: umidade relativa do ar, temperatura e pressão atmosférica influem significativamente na porcentagem de rainhas nascidas (Couto, 1991; Yodava, 1971 e Yodava e Smith, 1971 citados por Teixeira, 1993). Nooman e Kolmes (1989) pesquisaram o reconhecimento do parentesco da cria de operárias e de rainhas, por abelhas de duas patrilíneas de A.mellifera, e detectaram maior predisposição no cuidado dos ovos e larvas da mesma patrilínea. A rainha alimenta-se com geleia real tanto na fase larval como adulta. A geleia real é responsável pela diferenciação entre as castas, pelo desenvolvimento do aparelho reprodutivo da rainha, por sua longevidade e, segundo a quantidade de geleia real fornecida, às larvas apresentarão maior peso corporal ao nascer, maior número de ovaríolos e diâmetro de espermateca, quando comparadas com as mais leves (Souza et al., 2000). O objetivo deste trabalho foi avaliar a porcentagem de aceitação de larvas de diferentes grupos genéticos de A. mellifera, na produção de abelhas rainhas. Material e Métodos 1. Local O experimento foi desenvolvido no Apiário da Faculdade de Medicina Veterinária e Zootecnia, UNESP, Campus de Botucatu, com as seguintes coordenadas geográficas: 22° 49´de latitude sul e 48° 24´de longitude Oeste, com clima tipo Cfa segundo Köppem, e a coleta dos dados ocorreu no mês de julho de 2003. 2. Grupo Genético de Apis mellifera Foi considerado como grupo genético o conjunto de abelhas A. mellifera que pertencem a uma mesma subespécie de Apis (africanizadas ou italianas) e que podem ter diferentes graus de parentesco: meias irmãs, irmãs totais ou nenhum parentesco (Moura, comunicação pessoal). Para a realização deste trabalho foram estudados dois grupos genéticos:
3. Colmeias Mini Recrias Foram utilizadas 14 colmeias mini recrias. sendo:
Cada mini recria foi constituída por dois quadros com cria operculada e cerca de 6.000 abelhas aderentes, um quadro porta realeiras e um alimentador. 4. Colmeias Doadoras de Larvas Foram consideradas como colmeias doadoras de larvas, as 14 colmeias envolvidas no experimento, sendo sete do grupo genético italiano e sete do africanizado. 5. Tratamentos Os tratamentos realizados, para as colmeias mini recrias, foram:
Cada mini recria continha os três tratamentos nas transferências realizadas, nos diferentes períodos de orfandade. 6. Períodos de Orfandade Foram realizadas quatro transferências (períodos) para cada mini recria. Cada período foi constituído de 72 horas:
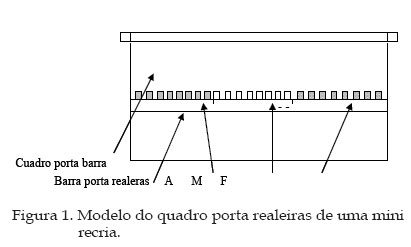
7. Localização das Larvas na Barra do Quadro Porta-Realeiras Para todos os tratamentos foram utilizasas os mesmos tipos de realeiras plásticas, cor laranja onde foram transferidas as larvas de 24 a 30 horas de vida, Logo apos de atrasferencia foram colocadas na barra superior do quadro porta realeiras, que continha os três tratamentos, num total de 24 realeiras em cada mini recria. Foram estabelecidos três locais (Figura 1):
8. Transferências As transferências foram feitas dentro do laboratório com temperatura média de 25oC e umidade relativa média de 70%. Os quadros porta realeiras foram colocados na parte central da colmeia mini recria. 9. Fatores Meteorológicos Os fatores meteorológicos medidos foram: temperaturas máxima, média e mínima (oC); umidade relativa do ar (%); pressão atmosférica (mmHg); precipitação pluviométrica (mm); velocidade do vento (km/dia) e pressão parcial de vapor (mmHg). As mediçoes metoerologicas foram feitas pela estação meteorologica da FMVZ Unesp. 10. Características Estudadas Foram avaliadas as seguintes características:
11. Delineamento Experimental e Métodos Estatísticos Foi utilizado o EIA: experimento inteiramente aleatorizado, e os dados foram submetidos à Análise de Variância não paramétrica, Kruskal-Wallis, e as médias que apresentaram diferenças significativas foram comprovadas pelo teste de Dunn (Curi, 1997). Resultados e Discussão 1. Porcentagem De Aceitação Das Larvas 1.1. Grupos genéticos e Tratamentos. A análise estatística dos resultados não mostrou diferença estatisticamente significativa entre o grupo genético italiano (58.78%) e o africanizado (60.11%), quanto a porcentagem de aceitação das larvas, (P>0.70) (Quadro 1). Esses resultados são semelhantes aos obtidos por Couto (1991) e Toledo (1997) que não encontraram diferença significativa entre as italianas e africanizadas, mas são diferentes daqueles encontrados por Duran (1991) e Garcia (1992) que obtiveram maior porcentagem de aceitação nas italianas do que nas africanizadas. Quanto aos tratamentos realizados, as análises estatísticas dos dados não mostraram diferenças significativas entre os tratamentos (P>30), tratamentos 1 (T1: 56.25%), tratamento 2 (T2: 63.39%) e o tratamento 3 (T3:58.64%), para os dois grupos genéticos. Esses resultados não concordam com os dados encontrados por Nooman e Kolmes (1989) e Page e Peng (2001) que observaram existir um reconhecimento das larvas provenientes da mesma matrilínea ou patrilínea, pelas operárias responsáveis pelos cuidados às larvas. Quadro 1. Porcentagem de aceitação das larvas, 72 horas após a transferência para as realeiras. nos dois grupos genéticos e nos três tratamentos
AFR: grupo genético africanizado; ITA: grupo genético
italiano; T1: tratamento 1: larvas da mesma matrilínea; T2: tratamento
2: larvas do mesmo grupo genético; T3: tratamento 3: larvas de
grupo genético diferente 1.2. Localização das realeiras na barra do quadro porta realeiras Observou-se que não houve diferença estatisticamente significativa entre os três locais (P>10), apesar da aceitação das larvas no local M ter sido 7.7% maior que a do local F e 15.8% maior que a do local A (Quadro 2). Esses resultados concordam parcialmente com os de Garcia (1992) que verificou menor aceitação, nas realeiras mais próximas do alvado, provavelmente devido a deficiência na termorregulação na região dessas realeiras. 1.3. Períodos de orfandade. Os resultados e a análise estatística mostraram que não houve diferença significativa entre os períodos (P>0.40), mas no período 1 (PO1) foi encontrada a maior aceitação (62.50%) e no período 4 (PO4) a menor (53.85%), mostrando uma tendência de diminuição da aceitação de acordo com o número de horas de orfandade. (Quadro 3) Quadro 2. Porcentagem de aceitação das larvas, 72 horas após a transferência para as realeiras nos três locais na barra do quadro porta realeiras
A: localização mais próxima ao alvado na barra do quadro
porta realeiras; M: localização na parte central; F: localização
na parte
mais distante do alvado 1.4. Postura de Operárias. Como as mini recrias não receberam reforço de cria durante todo o experimento, foram detectadas posturas de operárias a partir do período 3, em cinco mini recrias do grupo africanizado, quando as abelhas estavam com aproximadamente 11 dias de orfandade. Contudo, o grupo italiano só apresentou postura a partir do período 4 e em apenas duas mini recrias, quando as abelhas estavam com aproximadamente 14 dias de orfandade. Terada et al.(1975) e Kulencevic e Rothenbuhler (1982), citados por Garcia (1992) encontraram abelhas operárias mais velhas com comportamento de postura e que o componente genético era importante na determinação desta característica. Quadro 3. Porcentagem de aceitação das larvas, 72 horas após a transferência para as realeiras nos quatro períodos de orfandade
PO1: período 1, de 73 a 144 horas de orfandade; PO2: período
2; de 145 a 216 horas de orfandade; PO3: período 3, de 217 a 288
horas de orfandade; PO4: Período 4, de 289 a 360 horas de orfandade Assim, em mini recrias sem rainha, há necessidade de acréscimo de novas abelhas nutrizes, nove dias após a montagem das mini recrias para o grupo genético africanizado e depois de 12 dias para o grupo italiano. (Quadro 4) 2. Peso da galeia real 2.1. Grupos genéticos e Tratamentos. Quanto aos grupos genéticos, não foi encontrada diferença estatisticamente significativa entre eles (P>30), ou seja que a produção média de geleia real, por realeira, é semelhante nos grupos genéticos italiano (136.32 mg) e africanizado (117.63 mg). (Quadro 5) Provavelmente, o fato de obter-se menor quantidade de geleia real nas africanizadas, apesar de não ser significativa, seja devido ao ciclo de desenvolvimento mais curto que o das italianas (Rocha. 1999). Esses resultados concordam parcialmente com os encontrados por Garcia (1992) em que apesar das abelhas italianas produzirem mais geleia real, depositam-na mais lentamente nas realeiras, que as africanizadas e que a extração de geleia real das italianas deveria ser efetuada 72 horas após a transferência da larva, enquanto que para as africanizadas, seriam suficientes 69 horas. Quadro 4. Presença de postura de operárias em relação aos quatro períodos de orfandade
AFR: grupo genético africanizado; ITA: grupo genético italiano PO1: período 1, de 73 a 144 horas de orfandade; PO2: período 2, de 145 a 216 horas de orfandade; PO3: período 3, de 217 a 288 horas de orfandade; PO4: Período 4, de 289 a 360 horas de orfandade Quadro 5. Peso da geleia real (mg), 72 horas após a transferência para as realeiras nos dois grupos genéticos e três tratamentos
AFR: grupo genético africanizado; ITA: grupo genético
italiano; T1: tratamento 1: larvas da mesma matrilínea; T2: tratamento
2: larvas do mesmo grupo genético; T3: tratamento 3: larvas de
grupo genético diferente Quanto aos tratamentos, não foram encontradas diferenças estatisticamente significativas entre eles, (P>10). Consequentemente, para a produção de geleia real poderiam ser utilizadas larvas provenientes da mesma colônia ou matrilínea, do mesmo grupo genético ou de grupo genético diferente. (Quadro 5) 2.2. Grupos genéticos e Períodos de orfandade. A produção da geleia real, por realeira, dentro dos períodos de orfandade, verificou-se que houve diferença estatisticamente significativa entre os períodos de orfandade (P<0.01), sendo que os 1 e 2 não diferiram entre si e foram maiores significativamente que os 3 e 4, que também não diferiram entre si. Observou-se, também, que os dois grupos genéticos apresentaram produções decrescentes, mas no africanizado, este decréscimo foi mais acentuado, inclusive no período 4 (PO4) houve diferença significativa entre os dois grupos genéticos (P<0.05), apesar das médias não apresentarem diferença significativa (P>30). Encontrou-se que o grupo genético italiano manteve uma produção maior que a do africanizado, sendo que as italianas produziram 0.21%, 7.50%, 19.60% e 37.12% a mais que as africanizadas; para os períodos 1, 2, 3 e 4, respectivamente (Quadro 6). A diminuição da produção de geleia real, nos períodos 3 e 4, pode ter ocorrido pelo decréscimo da capacidade produtiva das abelhas nutrizes no decorrer do experimento, pois não houve reposição da população nas mini recrias, durante todo o experimento, além das condições climáticas mais adversas terem ocorrido nestes mesmos períodos. Couto (1991) encontrou melhores produções de geleia real e rainhas nos meses de dezembro a fevereiro, com redução gradativa dessas produções até junho. Esses resultados concordam, em parte, com os de Garcia (1992) que encontrou diferença significativa entre a produção de geleia real das abelhas africanizadas e italianas, sendo maior nas italianas e também com os de Couto (1991) que não encontrou diferença significativa entre italianas e africanizadas, mas que as italianas mostraram uma tendência a produzirem menos que as africanizadas, mas, não concordam com aqueles resultados coletados por Toledo (1997), que obteve maior produção para as africanizadas. 2.3. Localização das realeiras na barra do quadro porta realeiras e Períodos de orfandade. Quadro 6. Peso médio da geleia real (mg), 72 horas após a transferência para as realeiras nos quatro períodos e nos dois grupos genéticos
PO1: período 1, de 73 a 144 horas de orfandade; PO2: período
2; de 145 a 216 horas de orfandade; PO3: período 3, de 217 a 288
horas de orfandade; PO4: Período 4, de 289 a 360 horas de
orfandade; AFR: grupo genético africanizado; ITA: grupo genético
italiano Observou-se que houve diferença estatisticamente significativa entre os três locais (P<0.05), A, M e F quanto a produção meia (mg) por realeira, apresentando o local M maior produção do que os locais A e F que foram menores, sendo que os dois últimos não diferiram entre si (Quadro 7). Esses dados são concordantes com os de Garcia (1992) que registrou maior produção nas realeiras do meio. 3. Peso das larvas 3.1. Grupos genéticos e Tratamentos. Quadro 7. Peso da geleia real (mg), 72 horas após a transferência para as realeiras nos três locais na barra do quadro porta realeiras e nos quatro períodos
PO1: período 1, de 73 a 144 horas de orfandade; PO2: período 2; de 145 a 216 horas de orfandade; PO3: período 3, de 217 a 288 horas de orfandade; PO4: Período 4, de 289 a 360 horas de orfandade; A: localização mais próxima ao alvado na barra do quadro porta realeiras; M: localização na parte central; F: localização na parte mais distante do alvado Efeito do local: H=6.20; P<0.05 Letras maiúsculas diferentes indicam diferenças significativas entre locais (P<0.05) Não foram encontradas diferenças estatisticamente significativas no peso médio das larvas entre os grupos africanizado e italiano (P>30), apesar de se notar uma tendência das larvas africanizadas a terem maior peso que as italianas, nesta fase de desenvolvimento (Quadro 8). Esses resultados concordam com Rocha (1999), que trabalhando com a massa ponderal estimada das operárias de abelhas africanizadas, italianas e cárnicas, durante o ciclo de desenvolvimento, verificou que a massa ponderal das africanizadas foi superior às outras, no mesmo período de desenvolvimento em que se encontravam as larvas deste experimento. Foram encontradas diferenças estatisticamente significativas dos pesos das larvas entre tratamentos (P<0.01), sendo que as larvas do tratamento T2 foram significativamente maiores que as do T1 e T3, que não diferiram entre si. 3.2. Localização das realeiras na barra do quadro porta realeiras. Quando analisado o efeito dos locais no peso médio das larvas na barra do quadro porta realeiras, foram encontradas diferenças estatisticamente significativas entre eles (P<0.01), sendo que as larvas localizadas no meio (M) foram mais pesadas que as localizadas na posição F e A (Quadro 9). O maior peso das larvas do local M pode ser em decorrência da maior facilidade em manter a temperatura e a alimentação quando as condições externas e internas são adversas, como as ocorridas durante o experimento, pelas temperaturas baixas, chuva e ao baixo número de abelhas nutrizes das mini recrias, quando comparadas com outros experimentos, onde o número de abelhas por cada mini recria era aproximadamente 7000 a 8400 nutrizes (Garcia. 1992). Quadro 8. Peso da larva (mg), 72 horas após a transferência para as realeiras nos dois grupos genéticos e nos três tratamentos
AFR: grupo genético africanizado; ITA: grupo genético
italiano; T1: tratamento 1: larvas da mesma matrilínea; T2: tratamento
2: larvas do mesmo grupo genético; T3: tratamento 3: larvas de
grupo genético diferente Quadro 9. Peso da larva (mg), após 72 horas da transferência para as realeiras nos três locais na barra do quadro porta realeiras
A: localização mais próxima ao alvado na barra do quadro
porta realeiras; M: localização na parte central; F: localização
na parte
mais distante do alvado 4. Coeficientes de correlação de Pearson entre a porcentagem de aceitação das larvas, peso da geleia real e peso da larva. 4.1. Resultados Totais. Foram encontrados coeficientes de correlação positivos e altamente significativos entre: a porcentagem de aceitação das larvas e a produção da geleia real (r: 0.65, P<0.01); a porcentagem de aceitação e peso da larva (r: 0.52, P<0.01) e a produção da geleia real por realeira e o peso da larva (r: 0.59, P<0.01), (Quadro 10). Esses resultados indicam que quando maior é a porcentagem de aceitação, maior será a produção de geleia real e maior o peso da larva aceita, concordando com os resultados obtidos por Duran (1991) e Garcia (1992); mas discordando dos obtidos por Couto (1991). 4.2. Grupos genéticos. Quadro 10. Coeficientes de correlação de Pearson entre as variáveis: porcentagem de aceitação das larvas (%AC), peso da geleia real por realeira (PGR), peso da larva (PL), três tratamentos (T1.T2.T3), três locais do quadro porta realeiras (A.M.F) e dois grupos genéticos (AFR. ITA)
*significativo para P<0.05 **significativo para P<0.01 Foram encontrados coeficientes de correlação positivos e altamente significativos (P<0.01) entre: a porcentagem de aceitação das larvas e a produção da geleia real; entre a porcentagem de aceitação e peso da larva e, entre a produção da geleia real por realeira e o peso da larva para os dois grupos genéticos. Estes resultados indicam que, para os grupos genéticos africanizado e italiano, quando maior for a porcentagem de aceitação, maior será a produção da geleia real por realeira e maior o peso da larva. (Quadro 10) 4.3. Tratamentos. Foram encontrados coeficientes de correlação positivo e significativo (r :0,65; P<0.05) entre a porcentagem de aceitação das larvas e a produção da geleia real para os tratamentos T1, T2 e T3; entre a porcentagem de aceitação e o peso da larva para os tratamentos T1 e T2; e entre o peso da geleia real por realeira e o peso da larva nos tratamentos T1, T2 e T3, (Quadro 10). Esses resultados mostram que quanto maior for a quantidade de geleia real por realeira, maior será o peso da larva, que refletirá na produção de rainhas de maior peso, com maior volume da espermateca, maior número de ovaríolos e consequentemente maior capacidade de postura e vida útil, valendo tanto para as larvas provenientes da mesma matrilínea, ou mesmo grupo genético ou de outro grupo genético. Esses resultados concordam com os obtidos por Souza et al., (2000), que verificaram maior peso ao nascimento em rainhas com realeiras de maior volume. 4.4. Localização das realeiras na barra do quadro porta realeiras. Foram encontrados coeficientes de correlação positivo e significativo (r: 0,52; P<0.05) entre a porcentagem de aceitação das larvas e a produção da geleia real para os Locais A, M e F; entre a porcentagem de aceitação e a produção da larva para os Locais A e F; e entre a produção da geleia real por realeira e a produção da larva nos Locais A e F, (Quadro 10). Estes resultados indicam que nos locais extremos, A e F, a porcentagem de aceitação, produção da geleia real e o peso da larva são mais dependentes uns dos outros e, provavelmente, mais susceptíveis as mudanças no microclima da colmeia. Já, no local M, as condições ambientais internas da mini recria são mais estáveis. Estes resultados não concordam com os de Garcia (1992) que não encontrou relação entre a produção de geleia real por realeira e sua posição. 5. Fatores meteorológicos Verificou-se que apenas a precipitação pluviométrica correlacionou-se negativamente com a porcentagem de aceitação das larvas de um modo geral (r:-0,23; P<0,5), ou seja que quando há o aumento da chuva, há uma diminuição da porcentagem de aceitação. Somente o grupo genético africanizado sofreu influência significativa e negativa desse fator meteorológico, isto não ocorreu para o grupo italiano e para os diferentes locais das realeiras (r:0,01; P>0,01). Os outros fatores meteorológicos estudados não apresentaram influência na porcentagem de aceitação das larvas (Quadro 11). Portanto esses resultados não concordam com os encontrados por Duran (1991) e Garcia (1992) que não acharam correlação significativa entre a porcentagem de aceitação das abelhas africanizadas com os fatores climáticos, e diferem daqueles obtidos por Couto (1991) que achou correlação positiva entre o número de realeiras aceitas e a temperatura e, negativa com umidade relativa, nas abelhas italianas, mas não obteve correlação significativa para as africanizadas. Discordam, também, de Rinderer et al., (1985), citado por Couto (1991), que afirmou que as africanizadas são mais adaptadas que as européias às condições úmidas, e que, portanto, em ambientes úmidos, apresentam superioridade na produção de geleia real. Segundo Couto (1991) existem controvérsias na pouca bibliografia existente, sobre a ação do meio ambiente na aceitação das realeiras. Para Vuillaume (1957), citado por Couto (1991), a chuva reduz a produção de rainhas e para Allen (1965), também citado por Couto 1991, as condições climáticas desfavoráveis aumentam o número de operárias adultas no interior da colmeia, aumentando a aceitação das larvas. Para Mitev (1967), citado por Toledo (1997), a porcentagem de aceitação depende das condições climáticas, fluxo de néctar e número de larvas transferidas. Já para Corbella (1993), também citado por Toledo (1997), existe maior influência do estado interno da colônia do que das condições climáticas na porcentagem de aceitação. Quadro 11. Coeficientes de correlação de Pearson entre a variável: porcentagem de aceitação das larva (%AC) nos três locais da barra do quadro porta realeiras (A, M, F) e dois grupos genéticos (AFR, ITA) e os fatores meteorológicos
*significativo para P<0.05 **significativo para P<0.01 Verificou-se que a produção da geleia real no grupo genético africanizado apresentou coeficientes de correlação significativos com todos os fatores meteorológicos estudados, positivos com as temperaturas mínima, média e máxima e pressão de vapor e negativo com a umidade relativa, e o grupo genético italiano apresentou coeficiente de correlação significativo e negativo apenas com a umidade relativa. Provavelmente, os fatores meteorológicos afetaram menos ao grupo genético italiano quanto a produção da geleia real, do que ao grupo africanizado. Em relação aos locais onde estavam colocadas as realeiras na barra do quadro porta realeiras, o local A apresentou coeficientes de correlação significativos com todos os fatores meteorológicos estudados, sendo positivos com as temperaturas mínima, média e máxima, e com a pressão de vapor, e negativo com a umidade relativa do ar. Quanto ao local M, todos os coeficientes de correlação entre a produção da geleia real e os fatores meteorológicos não foram significativos (P>0,05). Quanto ao local F, o coeficiente de correlação foi significativo e positivo ( r:0,41; P>0,05) entre a produção da geleia real e a temperatura máxima e negativa com a umidade relativa (r: -0,43; P<0,01). Portanto, os fatores meteorológicos influenciaram significativamente na produção de geleia real para o local A, não influíram para o local M e influíram parcialmente na produção de geleia real para o local F. (Quadro 12) Considerações finais Para a produção de rainha, a utilização de larvas, sejam provenientes da mesma matrilínea, do mesmo grupo genético ou grupo genético diferente, não influenciou significativamente a porcentagem de aceitação das larvas Quadro 12. Coeficientes de correlação de Pearson entre a variável: peso da geleia real por realeira (PGR), três tratamentos (T1,T2,T3), três locais das realeiras na barra do quadro porta realeiras (A, M, F) e dois grupos genéticos (AFR, ITA) e os fatores meteorológicos
*significativo para P<0.05 **significativo para P<0.01 Para a produção de rainha, a localização das larvas na barra do quadro porta realeiras não influenciou significativamente, a porcentagem de aceitação das larvas. Em mini recrias sem rainha, deveria haver acréscimos de novas abelhas nutrizes, nove dias após a montagem das mini recrias para o grupo genético africanizado e, de 12 dias para o grupo italiano. Para a produção de rainhas, os grupos genéticos africanizado e italiano não apresentaram diferenças significativas na produção da geleia real (mg), por realeira. quando foram utilizadas mini recrias. No inverno, os fatores meteorológicos: temperaturas mínima e máxima, umidade relativa do ar e pressão de vapor afetaram significativamente a produção da geleia real produzida pelas abelhas africanizadas. No inverno, as mini recrias italianas foram menos susceptíveis às mudanças dos fatores meteorológicos do que as africanizadas, sendo influenciada significativamente apenas pela umidade relativa do ar. No inverno, os fatores meteorológicos: temperaturas mínima e máxima, umidade relativa do ar e pressão de vapor afetaram significativamente a produção da geleia real por realeira, quando estas estavam localizadas em local mais próximo do alvado. No inverno, os fatores meteorológicos: temperatura máxima e a umidade relativa do ar influíram significativamente a produção da geleia real por realeira, quando estas estavam localizadas em local mais afastado do alvado. No inverno, nenhum dos fatores meteorológicos estudados afetou significativamente a produção da geleia real por realeira, quando estas estavam localizadas no meio da barra do quadro porta realeiras. Para produzir uma rainha de boa qualidade no inverno de Botucatu, SP, as realeiras deveriam ser localizadas na parte central do quadro porta realeiras. Conclusões A partir dos resultados obtidos neste experimento, pode-se concluir que:
Literatura Citada
© 2006 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la06005f1.jpg] [la06005t1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}