 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
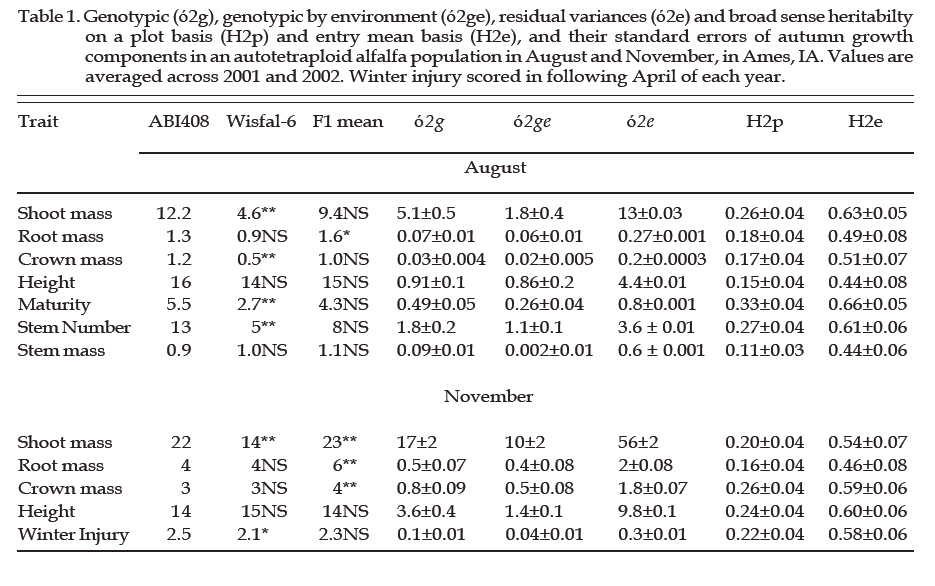
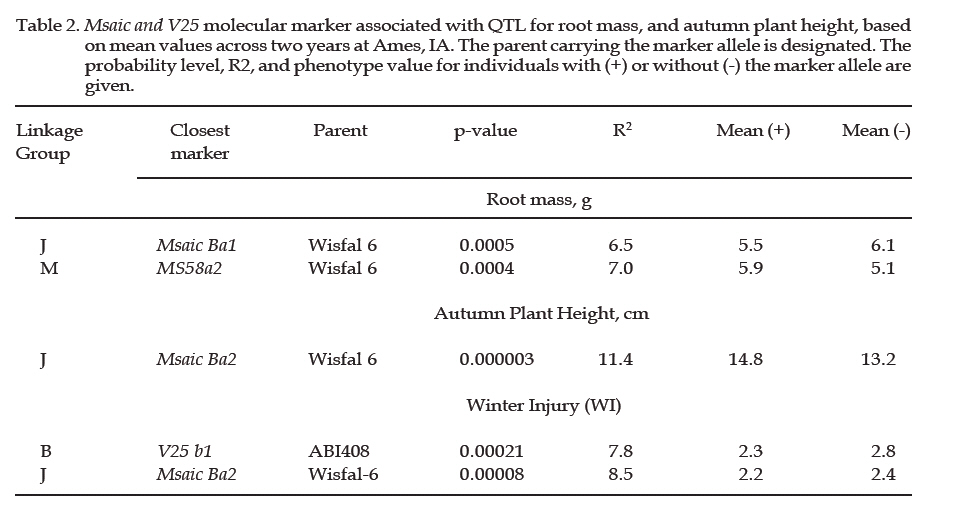
Archivos Latinoamericanos de Produccion Animal, Vol. 15, No. 2, 2007, pp. 70-73 A candidate gene-marker approach to improve coldhardiness in alfalfa (Medicago sativa) B. Alarcón Z1 and E. B. Charlie2 1Universidad Autónoma Chapingo, Chapingo, Estado de México, México. 56230. E-mail: b_alarcon_zuniga@yahoo.com.mx 2Agronomy Department. Iowa State University, Ames, IA, USA Code Number: la07011 ABSTRACT In winter hardy alfalfa cultivars, cold acclimation occurs prior to the onset of freezing temperatures, and normally is accompanied with a series of genetic, metabolic and morphological adjustments. We investigated the association between three alleles derived from cold-regulated genes (Msaic&V25) and the metabolic and morphological winter hardiness traits, in an F1 segregating population between the cross of M. sativa ssp. sativa (ABI408) and falcata (Wisfal-6). Transgressive segregation was observed in all traits. Broad sense heritability ranged from low for stem weight to high for shoot mass. The presence of allele a1 of Msaic B, a cold-regulated gene (COR), was positively associated with plant autumn height but negatively associated with root mass in the Wisfal-6 parent and not association to taproot metabolites content; it explained up to 11.4% of the winter hardiness variation in the segregating population. V25b1, a COR gene, was associated to winter injury, explaining 7.8% of the phenotypic variation. Both RFLP probes can be used as markers for cold hardiness breeding scheme. Our results also suggest that winter injury and autumn biomass are controlled by different loci in this population. Key words: alfalfa; winter hardiness; COR genes/markers Titulo faltaRESUMEN En variedades de alfalfa tolerantes al frío, el periodo de aclimatación ocurre previo al inicio de la presencia de temperaturas de congelamiento invernales, y este cambio es acompañado con una serie de ajustes genéticos, metabólicos y morfológicos en las plantas. En este experimento, se investigó la asociación entre 3 alelos derivados de genes regulados por frío (genes COR: Msaic&V25) y los componentes de crecimiento en tallos, hoja, raíz, número de tallos y altura de la planta, en una población segregante F1 de la cruza de M. sativa ssp. sativa (ABI408) y falcata (Wisfal-6). Se observó segregación transgresiva en todos los caracteres morfológicos de la población, con una heredabilidad en sentido amplio de baja para peso de tallo a alta para biomasa total de ramas. La presencia del alelo a1 de Msaic B, un gen regulado por frío (COR), fue positivamente asociado con la altura de la planta en el otoño pero negativamente asociado con la biomasa de raíces en el progenitor Wisfal-6, y no se encontró una asociación con el contenido de metabolitos en raíces superiores. Este alelo explicó hasta un 11.4% de la variación fenotípica en la tolerancia a invierno en la población segregante. V25b1, un gen COR, se asoció con el daño invernal, explicando 7.8% de la variación fenotípica. Ambas sondas de RFLP pueden ser empleadas como marcadores para mejoramiento genético en la tolerancia a invierno. Nuestros resultados también sugieren que el daño invernal y la acumulación de biomasa en otoño son controlados por loci diferentes en esta población. Palabras clave: alfalfa; tolerancia a invierno; genes COR/ marcadores IntroductionGenetic evidence suggests that induction of cold hardening in alfalfa via autumn acclimation is a quantitative trait controlled by a number of additive genes (Brummer, 2004; Weishaar et al., 2005), leading to differential gene expression. A growing number of genes and cDNAs that are induced by cold have been cloned in cold acclimated seedlings (Mohapatra et al., 1988) or crowns (Laberge et al., 1993). Msaic cDNA clone is a cold and growth regulated gene in alfalfa. Msaic encodes a putative Gly-rich protein (38%), which contains many repeated motifs, sharing homology in the range of 68 to 88% (amino acid identity) with isolated environmental stress- and ABA-regulated putative proteins from alfalfa (Luo et al., 1991, 1992), and represent a new member of the cold-regulated (COR) gene family. Development of Restricted Fragment Length Polymorphism (RFLP) probes and Simple Sequence Repeat-Expressed Sequence Tag (SSR-EST) gene-selective primer sequence has been a common practice in marker assisted breeding programs, and they are widely used to identify target genes, quantitative trait loci (QTL), number and frequency of allele among populations (Alarcon et al., 2004). Herein, we propose a novel breeding scheme to target cold regulated genes in alfalfa, and its association to both morphological and metabolic winter hardiness traits, as a marker breeding approach. Material and MethodsPlant material: ABI408 (Medicago sativa ssp. sativa) and Wisfal-6 (M. sativa ssp. falcata) genotypes were crossed to form an F1 population of 200 individuals segregating for yield, winter injury and autumn growth (Brummer et al., 2000). The 200 F1 individuals, two parents and eight checks were clonally propagated by stem cuttings in the greenhouse at 27-32ºC, watered as required, fertilized with 20-30-10 three times, and at 4 weeks-old field transplanted in Ames, Iowa on June 1, 2001, and on May 15, 2002. The plot design in both years was a 14 by 15 quadruple alpha lattice design with 3 replications and 3 destructive harvest dates defined as blocks (August, November and April). Five plants per genotype were space planted at 16 cm apart within a plot in each replication with rows separated by 80 cm. Morphological and chemical analysis: Plant height was measured in late August and early November, that each plant was dug, washed in water, and taproots, crowns and shoots were separated and biomass recorded. The following April, winter injury was scored following McCaslin and Woodward’s method (1995), which included the digging of the complete plant and visual scoring on a scale of 1= no injury, all plant symmetrical with equal length, up to 5 = all dead plants. Soluble carbohydrates, total protein, amino-N groups and lipids on taproots were analyzed. COR gene-marker development: Restricted Fragment Length Polymorphism (RFLP) probes were developed from the RNA extraction and reversible transcripted to cDNA (GenBank/EMBL Acc. L03708), and surveyed to parental and F1 individual blots. In addition, Medicago SSR primer pair sequence designed by software Primer3 v.1.2 of HHMIBR & NIH-NGH-Genome, Wisconsin, and AFLP (Amplified Fragment Length Polymorhism) EcoR1 y MseI primer combinations were screened to the parental and F1 segregating population. Statistical analysis: Analysis of variance of each trait was done in each harvest date and combining across years with all effects in the model considered as random (Lynch and Walsh, 1998). Variancecovariance matrix components, heritability and single dose restriction fragments were carried out (Falconer and Mackay, 1996). Single marker analysis was performed to estimate association between traits and marker loci with a probability level of P<0.001. The mean for each trait as marker was considered as present (+) or absent (-) to estimate phenotypic effect of the marker alleles. Results and DiscussionThere was highly significant difference between parents, but no difference to F1 population, in both harvest dates for all morphological traits. Therefore, additive effect was assumed for all traits, except root mass. The progeny showed transgressive segregation for all traits. Genotypic (ó2g), genotypic x environment (ó2ge), and residual variances (ó2e) were also significant for all traits. The broad sense heritabilities on a plot and entry mean basis ranged from low for stem weight to high for shoot mass, similar as reported by Dhont et al., (2003) Table 1. Msaic Ba1 and MsaicBa2 were identified into linkage group J and M, associated to roots mass and autumn plant height. No association was identified between these two probes and metabolic components (data not shown), assuming a strong relationship to alfalfa autumn growth and fall dormancy (Laberge et al., 1993). MsaicBa2 interpreted 11.4% and 8.5% of the phenotypic variation for autumn plant height and winter injury, respectively. V25 b1, a COR gene related to soluble carbohydrates, explained 7.5% of the WI Table 2. ConclusionsWe have demonstrated that we can identify loci involved in important physiological and biochemical pathways underlying complex agronomic traits. Further analysis will focus on identifying the precise regions of both COR genes and sequence homology to other populations and forage species. Literature Cited
© 2007 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la07011t2.jpg] [la07011t1.jpg] |
| |||||||||

{kind=link}
{kind=link}