 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
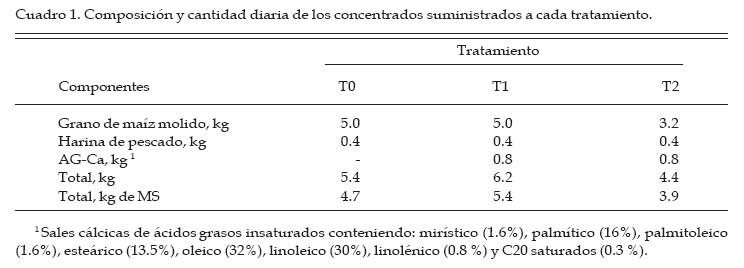
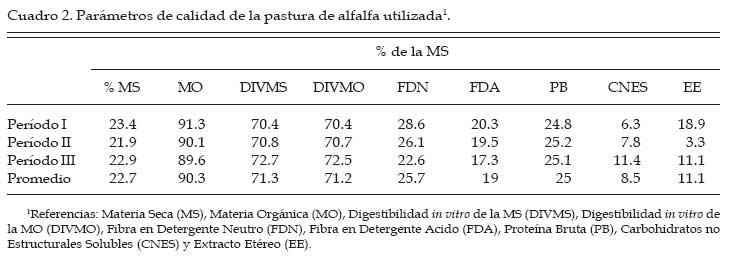
Archivos Latinoamericanos de Produccion Animal, Vol. 15, No. 3, 2007, pp. 88-99 Efectos de la suplementación con sales cálcicas de ácidos grasos insaturados sobre la digestión ruminal en vacas lecheras en pastoreo Effects of calcium salts of unsaturated fatty acids supplementation on ruminal digestion by grazing dairy cows G. A. Gagliostro1 y G. F. Schroeder2 1Instituto Nacional de Tecnología Agropecuaria (INTA). Estación Experimental Balcarce, CC 276, (7620), Balcarce, Argentina Code Number: la07014 ABSTRACT The objective of this study was to evaluate the effects of calcium salts of unsaturated fatty acids (Ca-FA) supplementation on rumen digestion by dairy cows grazing high-quality pastures. Three cows with cannulas in the rumen were allocated to three dietary treatments in a 3×3 Latin square design. The cows grazed an alfalfa pasture with a herbage allowance of 30 kg dry matter (DM)/cow. Mean pasture quality was: 23% DM, 71% DM in vitro digestibility, 25% crude protein (CP), and 26% neutral detergent fiber (NDF). The treatments consisted in the supplementation with: T0 (5 kg of ground corn plus 0.4 kg of fishmeal), T1 (similar to T0 plus de addition of 0.8 kg de Ca-FA), and T2 (3.2 kg of ground corn, 0.4 kg of fishmeal, and 0.8 kg of Ca-FA replacing a similar part of the corn on an energy basis). The Ca-FA contained 65% of unsaturated and 31% of saturated FA. Total volatile fatty acids (105 mmoles/L), acetato to propionate ratio (3.0), and ruminal pH (5.8) were not affected by Ca-FA supplementation. Isobutyrate concentration in the rumen was increased in T0 (P<0.01). Parameters of in situ ruminal degradation of DM, NDF, and CP of the pasture were similar among treatments. Neither milk production nor milk composition was affected by the treatments. Overall, the results indicated that unsaturated FA supplementation in the form of Ca-FA avoided negative effects on ruminal pasture digestion, even when ruminal pH was relative low. Key words: fat, supplementation, rumen digestion, grazing RESUMEN El objetivo del estudio fue evaluar los efectos de la suplementación sales cálcicas de ácidos grasos insaturados (AG-Ca) sobre la degradación in situ del forraje en vacas en pastoreo. Tres vacas multíparas con fístula ruminal fueron asignadas a tres tratamientos en un cuadrado latino (3×3). Los animales pastorearon pasturas de alfalfa con una asignación diaria de 30 kg materia seca (MS)/vaca y una calidad promedio de 23% MS, 71% digestibilidad in vitro de la MS (DIVMS), 25% proteína bruta (PB) y 26% fibra en detergente neutro (FDN). Los tratamientos fueron: T0 (5 kg de maíz molido y 0.4 kg de harina de pescado), T1 (idem T0 más 0.8 kg de AG-Ca) y T2 (3.2 kg de maíz molido, 0.4 kg de harina de pescado y 0.8 kg de AG-Ca en sustitución isoenergética del maíz). Las AG-Ca utilizadas contenían 65% de AG insaturados y un 31% de AG saturados. Las concentraciones totales de ácidos grasos volátiles (105 mmoles/L), la relación acético/propiónico (3.0) y el pH ruminal (5.8) no fueron afectados por la suplementación con AG-Ca. La concentración de isobutirato fue mayor en T0 (P<0.01). La degradación ruminal de la MS, FDN y PB del forraje no difirió entre tratamientos. Tampoco se detectaron diferencias en la producción y composición de la leche. Los resultados sugieren que la suplementación con lípidos bajo la forma de sales cálcicas de AG insaturados parece haber sido adecuada al no producir efectos negativos sobre la digestión ruminal, aun cuando el pH ruminal fue relativamente bajo. Palabras claves: lípidos, suplementación, digestión ruminal, pastoreo Introducción En condiciones pastoriles de alimentación, durante el primer tercio de lactancia, las vacas lecheras sufren un importante déficit energético (Bargo et al., 2003). Consecuentemente, se observa un incremento en la movilización de reservas corporales y, en muchos casos, una reducción en la performance productiva y reproductiva de los animales. En respuesta a ello, a fin de incrementar el consumo de energía, la suplementación con granos de cereales es la alternativa más difundida en dichos sistemas (Bargo et al., 2003; Schroeder et al., 2004). Sin embargo, el creciente aumento de la proporción de granos en la dieta puede traer otros problemas asociados, tales como acidosis ruminal, disminución de la digestibilidad de la fibra, cambios en la relación acético/propiónico y disminución en el tenor graso de la leche (Bauman y Griinari, 2001). Frente a este problema surge la posibilidad del suministro de lípidos inertes a nivel ruminal, debido a su alta densidad energética (tres veces superior a la de los granos de cereales) y por no ser fermentados en el rumen (Palmquist y Jenkins, 1980). Además, la suplementación con ácidos grasos (AG) insaturados, no solo incrementa la densidad energética de la dieta, sino que también aportaría precursores de AG insaturados y otros isómeros (i.e. ácido linoleico conjugados) de la leche de gran importancia para la salud humana (Bauman y Griinari, 2001; Schroeder et al., 2003). Sin embargo, si los lípidos no son completamente inertes a nivel ruminal, y sobre todo cuando los AG son insaturados, se pueden producir efectos negativos sobre la fermentación ruminal, con disminución en la digestibilidad de la fibra (Tamminga y Doreau, 1991; Jenkins, 1993). Estos efectos a nivel ruminal resultan predisponentes a una disminución del consumo, de la producción de leche y del tenor proteico de la misma (Palmquist y Jenkins, 1980). Con el fin de evitar estos efectos negativos en rumen se han desarrollado numerosas técnicas de protección de lípidos, siendo la formulación de sales cálcicas de ácidos grasos (AG-Ca) la mayormente utilizada (Gagliostro y Chilliard, 1992). Si bien la combinación con cationes Ca parece ser una forma eficaz para atenuar los efectos negativos de los AG libres en el rumen, la estabilidad de dichos compuestos depende del grado de insaturación de los AG que lo componen y del pH ruminal (Palmquist y Jenkins, 1980). El pKa de los AG-Ca a base de ácido esteárico (C18:0) es de 4.5, mientras que es de 5.6 para las AG-Ca conteniendo AG insaturados (Sukhija y Palmquist, 1990). En rumiantes pastoreando forrajes de alta calidad, el pH ruminal es normalmente más bajo que en condiciones de confinamiento con dietas basadas en forrajes conservados (Bargo et al., 2003). Las investigaciones previas que evaluaron AG insaturados bajo la forma de AG-Ca fueron conducidos en condiciones de confinamiento y en situaciones donde los valores de pH ruminal se encontraban entre 6.1 y 6.4 (Chalupa et al., 1986, Doreau et al., 1993). Resulta necesaria, entonces, la evaluación estos suplementos en condiciones de alimentación pastoril, donde el pH ruminal se encontraría cercano al pKa de las AG-Ca. Por lo tanto, el objetivo del presente estudio fue evaluar el uso de sales cálcicas de AG insaturados en vacas lecheras pastoreando pasturas de alta calidad sobre la digestión ruminal y la producción y composición de la leche. Materiales y Métodos Animales y Tratamientos experimentales El ensayo fue llevado a cabo en la Estación Experimental de Balcarce dell Instituto Nacional de Tecnología Agropecuaria (INTA), Argentina, durante los meses de octubre y noviembre. Se utilizaron tres vacas de raza Holando Argentino con 46 ± 12 días en lactancia, provistas de fístula ruminal permanente. Las vacas fueron asignadas a un cuadrado latino 3×3, con tres tratamientos y tres períodos experimentales de 18 días de duración cada uno (14 días de adaptación al tratamiento y 4 días de toma de datos). Las vacas pastorearon una pastura monofítica de alfalfa (Medicago sativa) en un sistema de pastoreo en franjas diarias. El área de cada franja fue ajustada diariamente a fin de asegurar una oferta diaria de forraje no menor a 30 kg de MS/vaca, nivel donde se alcanzarían los máximos consumos de forraje (Minson, 1990). Sobre esta base pastoril de alimentación se implementaron siguientes tres tratamientos (Cuadro 1):
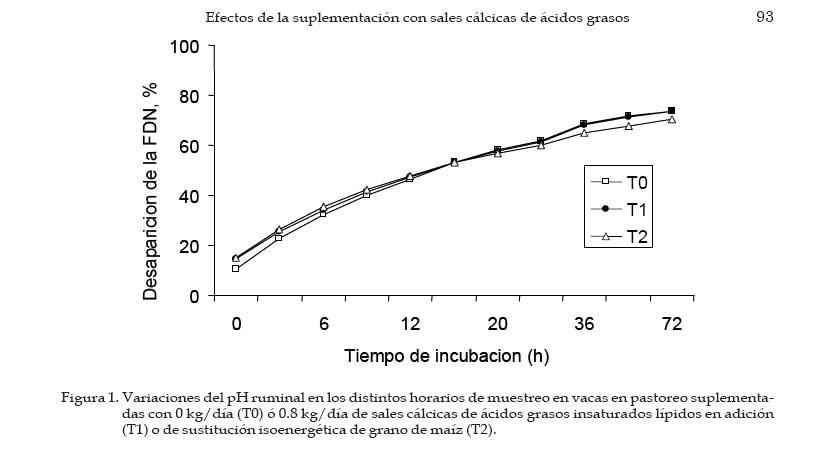
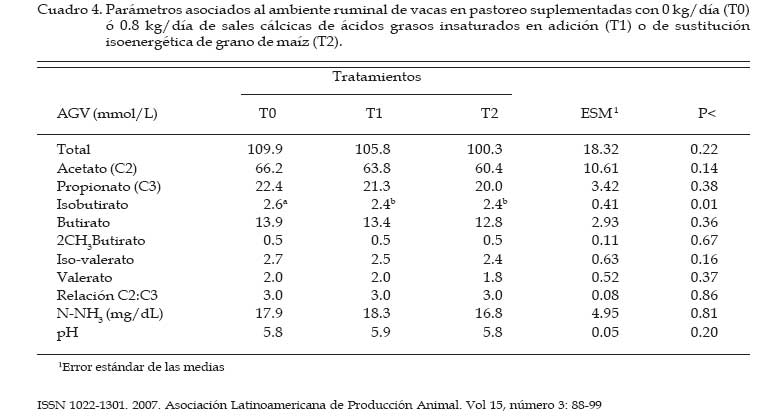
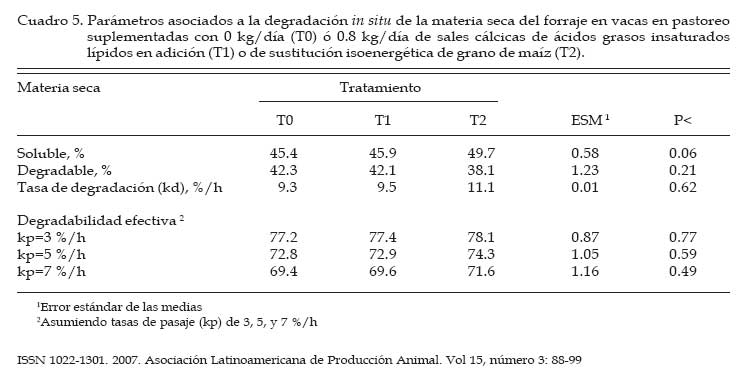
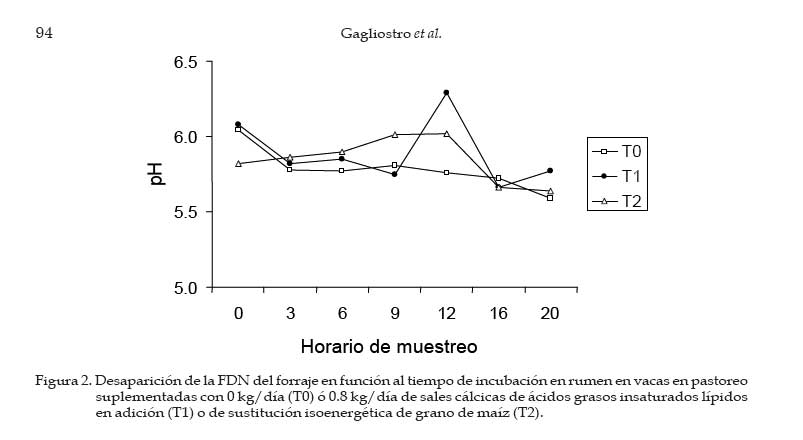
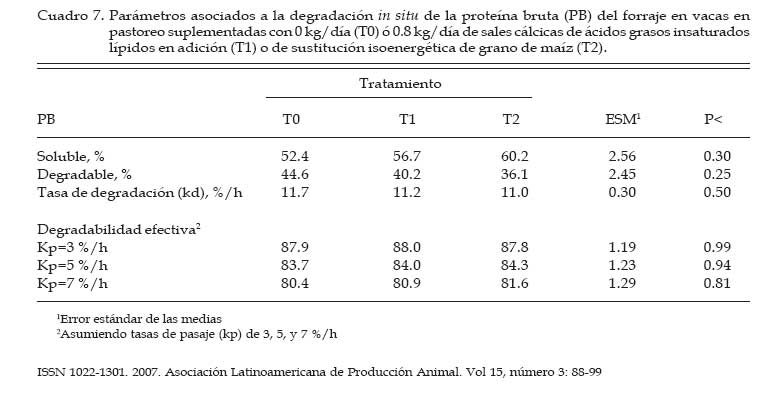
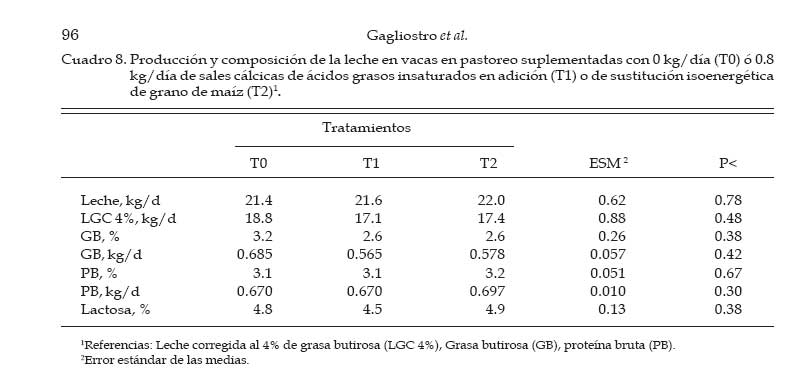
Los concentrados se suministraron en mitades durante cada turno de ordeño (06:00 y 18:00 hs). Las AG-Ca (Full Fat BP, Inagro, Buenos Aires, Argentina) contenían un 65% de AG insaturados y un 31% de AG saturados. El T1 se implementó con el objetivo de evaluar el efecto de las AG-Ca cuando la única fuente de variación respecto al tratamiento control (T0) fue el agregado de lípidos en el concentrado. En el T2 se reemplazó parte de la energía fermentescible en rumen (almidón aportado por el grano de maíz) por energía no fermentescible en rumen (AG-Ca). Por lo tanto, esos dos concentrados (T0 y T2) fueron formulados para aportar similar cantidad total de energía, pero bajo distintas formas químicas (carbohidratos y lípidos, respectivamente), lo que induce diferentes sitios y productos finales de la digestión. La dosis de AG-Ca (0.8 kg/vaca/día) fue calculada a fin de aportar un 13% de los requerimientos totales de energía metabolizable (EM) bajo la forma de lípidos, como fue recomendado por Palmquist y Jenkins (1980). El requerimiento total de EM fue calculado utilizando las ecuaciones propuestas por el NRC (2001) para una vaca promedio en inicio de lactancia de 580 kg de peso vivo y produciendo 25 kg de leche/día con 38.6 g/kg de grasa butirosa. Estos valores correspondieron al promedio de los datos obtenidos en el mismo rodeo experimental durante los primeros 30 días de la lactancia anterior (Salado et al., 2004). El valor energético de los lípidos (7.14 Mcal EM/kg MS) fue estimado a partir de su energía bruta (9.4 Mcal EB/kg MS) asumiendo una digestibilidad promedio de los AG del 80% (Palmquist y Jenkins, 1980) y un tenor de MS del 95%. Las AG-Ca utilizadas contenían un 85% de lípidos sobre base MS. Colección de datos experimentales La biomasa total de pastura fue determinada semanalmente mediante cortes a ras del suelo con tijera manual de un área delimitada por un marco de 0.1 m2, cortando una superficie total de 2 m2 en cada muestreo. La calidad del forraje y de los concentrados fue estimada al inicio de cada período experimental (Período I = 27/10; Período II = 15/11; Período III = 03/12) a partir de muestras extraídas en forma manual simulando el pastoreo selectivo que realizaban las vacas (hand-plucking). Sobre dichas muestras se determinó el contenido de MS (estufa a 80ºC durante 48 hs), materia orgánica (MO; mufla a 600ºC durante 4 hs), digestibilidad in vitro de la materia seca (DIVMS) y materia orgánica (DIVMO; Tilley y Terry, 1963), proteína bruta (PB; AOAC, 1990), fibra en detergente neutro (FDN) y ácido (FDA; AOAC, 1990), carbohidratos no estructurales solubles (CNES; AOAC, 1990) y extracto etéreo (EE; AOAC, 1990). Se realizaron mediciones de pH, concentración de nitrógeno amoniacal (N-NH3) y de ácidos grasos volátiles (AGV). Para ello se extrajeron muestras de licor ruminal a las 0, 3, 6, 9, 12, 16 y 20 hs de comenzado el muestreo (06:00 a.m.). El licor ruminal fue filtrado con tela tipo quesera e inmediatamente se midió el pH utilizando un pH-metro digital portátil (Orion portable pH meter 250A, Orion Research Inc., Boston, MA, USA). Una alícuota de 100 mL del licor filtrado fue trasvasada a botellas plásticas conteniendo 1 mL de ácido sulfúrico concentrado (H2SO4), y congeladas (-24°C) hasta ser analizadas. Las muestras fueron descongeladas a temperatura ambiente, centrifugadas a 10000 r.p.m. durante 20 min a 0°C, realizándose las determinaciones sobre el sobrenadante. Las concentraciones de N-NH3 fueron determinadas mediante lectura colorimétrica en autoanalizador de flujo continuo Technicon (Modelo 2). Las concentraciones de los AGV fueron determinadas por cromatografía gaseosa en un cromatógrafo Shimadzu GC-14, equipado con un integrador Shimadzu C-R6A, columna de vidrio de 2 m × 2 mm de diámetro interno, rellena con chromosorb AW, 10% polietilenglicol adipato y 3% H3PO4. Las temperaturas en la columna, el inyector y el detector fueron: 155°C, 185°C y 190°C, respectivamente. Las tasas de flujo empleadas para el carrier (N2), el hidrógeno (H2) y el aire cromatográfico fueron de: 50, 20 y 300 mL por min, respectivamente. El volumen inyectado fue de 1 mL de muestra. Para determinar la tasa y extensión de la desaparición de las diferentes fracciones del forraje se utilizó la técnica in situ (Mehrez y Ørskov, 1977). Se utilizaron 22 bolsas de dacrón (15.5 cm por 7.5 cm con un tamaño promedio de poros de 52 mm.) por vaca en cada período. En cada bolsa se colocó una muestra de forraje fresco conteniendo 5 g de MS. El forraje fue previamente picado a un tamaño de 1 cm. Las bolsas se introdujeron en el saco ventral del rumen, todas juntas a la hora 0 de muestreo (06:00 a.m.) y se fueron extrayendo por duplicado en once horarios de incubación en el rumen (0, 3, 6, 9, 12, 16, 20, 24, 36, 48 y 72 h). Cada bolsa fue lavada bajo corriente de agua fría hasta obtener un líquido incoloro eliminándose el material soluble y secadas en estufa (60ºC durante 48 h) y el residuo fue molido a 1 mm para las determinaciones de FDN y PB. La degradación potencial de la MS y de la PB del forraje se calculó utilizando la siguiente ecuación: P = S + D (1- e -kd.t ), donde, P = degradabilidad (%), S = fracción soluble (%), D = fracción potencialmente degradable (%), e = base del logaritmo natural, kd = tasa de degradación (%/h) y t = tiempo de incubación en rumen (horas). Los parámetros de dicha ecuación se calcularon mediante el método Marquardt para modelos no lineales del SAS (Schroeder et al., 2002). A partir de los resultados de degradabilidad potencial se calculó la degradabilidad efectiva asumiendo distintas tasas de pasaje (kp, 3, 5 y 7 %), mediante la ecuación propuesta por Ørskov y McDonald (1979) : E = S + [ D (kd / kd + kp)], donde, E = degradabilidad efectiva (%), S = fracción soluble, D = fracción degradable (%), kd = tasa de degradación (%/h) y kp = tasa de pasaje (%/h). Para el cálculo de la degradabilidad ruminal de la FDN se evaluaron dos modelos, con o sin incluir un tiempo de retardo (lag-time). Sobre la base de los errores mínimos cuadrados de cada modelo, y como no se evidenció la existencia de ningún tiempo de retardo, se procedió al análisis con un modelo similar al descripto para MS y PB como fue sugerido por Schroeder et al. (2003) para forrajes de alta calidad. La producción de leche fue diaria e individualmente determinada durante los últimos cuatro días de cada período experimental, tomando de cada vaca dos submuestras en ordeñes consecutivos (mañana y tarde) para luego confeccionar una muestra compuesta ponderada por la respectiva producción de leche. Sobre dichas muestras se determinó el contenido de grasa butirosa (GB), proteína bruta (PB) y lactosa (Foss 300, Milko-Scan; Foss Electric, Hillerod, Dinamarca). Análisis estadísticos Los datos fueron analizados mediante el procedimiento GLM (SAS, versión 6.12, 1996) según lo propuesto por Schroeder et al. (2002) en un diseño en cuadrado latino 3×3 utilizando el siguiente modelo: Yijk = m + Ai + Pj + Tk + eijkt, donde, Yijk = variable respuesta de la i-ésima vaca en el j-ésimo período correspondiente al k- ésimo tratamiento, m = media general, Ai = efecto de la i-ésima vaca, Pj = efecto del j-ésimo período experimental, Tk = efecto del k-ésimo tratamiento y eijk = error residual. Las variables ruminales (N-NH3, pH y AGV) fueron previamente analizadas en un diseño en cuadrado latino con mediciones repetidas en el tiempo utilizando el procedimiento MIXED del SAS a fin de evaluar el efecto del tratamiento, del horario de muestreo y la interacción entre ambos. Debido a que dicha interacción no resultó significativa (P>0.10) para ninguna de las variables analizadas, se promediaron los resultados de los diferentes horarios de muestreo y se analizó según el modelo anteriormente descripto. Se consideraron diferencias significativas si P<0.05 y tendencia cuando 0.05 > P < 0.10. Todas las comparaciones entre medias de tratamientos se realizaron mediante el test para medias ajustadas de Tukey- Kramer (Montgomery, 1991). Resultados Calidad de la pastura y concentrados experimentales La biomasa forrajera aérea promedio durante el estudio fue de 2764 ±379 kg MS/ha. En el Cuadro 2 se presenta la calidad nutritiva de la pastura de alfalfa utilizada durante cada uno de los períodos experimentales. La composición química de los concentrados se presentan en el Cuadro 3. La harina de pescado utilizada fue de alta calidad, con un contenido promedio de PB de 63.4% y un contenido de EE de 8.1%. Parámetros asociados al ambiente ruminal y digestión del forraje El efecto de la suplementacion con AG-CA sobre el ambiente ruminal se presenta en el Cuadro 4. La inclusión de AG-Ca en el concentrado ofrecido a vacas lecheras en pastoreo no modificó significativamente la concentración total de AGV, ni la relación C2:C3 sea en condiciones de adición (T1) como en condiciones de sustitución de almidón por lípidos (T2). Sólo la concentración de isobutirato presentó diferencias significativas entre tratamientos, resultando mayor en T0 (P<0.01). En la Figura 1 se presentan las variaciones del pH ruminal registradas a lo largo de 24 hs de medición. No se observaron efectos de la hora de muestreo (P<0.15) ni de la interacción entre la misma y los tratamientos (P<0.88). Las horas 0 y 12 de muestreo coinciden con las 6 y 18 hs, momento en el cual las vacas eran ordeñadas y se les suministraba el concentrado. Sólo en estos momentos se registraron valores de pH superiores a 6. La inclusión de lípidos en la ración no modificó significativamente las concentraciones promedio de N-NH3 (P<0.81), como puede observarse en el Cuadro 4. El efecto de la hora de muestreo resultó significativo (P<0.01) mientras que la interacción entre el horario de muestreo y los tratamientos resultó no significativa (P<0.58) Las concentraciones más altas de N-NH3 se observaron en la hora 6 de muestreo para T0 y T2 (34 y 21.9 mg/dL, respectivamente) y en la hora 9 para T1 (32.3 mg/dL), y los mínimos en las horas 3 y 12 (11.4 y 11.0 mg/dL, respectivamente) para todos los tratamientos. Los parámetros de desaparición ruminal de la MS de la pastura de alfalfa se presentan en el Cuadro 5. La fracción soluble tendió a ser mayor (P<0.06) en el T2, mientras que en el resto de los parámetros analizados no se detectaron diferencias significativas entre tratamientos (Cuadro 5). La degradabilidad potencial del forraje incubado fue similar (T0 = 87.7 %, T1 = 88.0 % y T2 = 87.8 %, P<0.96) entre los tratamientos. Los parámetros asociados a la desaparición ruminal de la FDN del forraje incubado en el rumen se presentan en el Cuadro 6. El mejor ajuste para el cálculo de los parámetros de cinética de desaparición de la FDN fue un modelo sin tiempo de retardo, tal como puede observarse en la Figura 2. No se detectaron diferencias estadísticamente significativas para ninguno de los parámetros analizados (Cuadro 6). La fracción potencialmente degradable de la FDN resultó elevada sin diferencias significativas entre tratamientos (T0 = 74.4%, T1 = 74.7% y T2 = 72.4%; P<0.70). Los parámetros asociados a la desaparición ruminal de la PB del forraje se presentan en el Cuadro 7. La inclusión de AG-Ca en los concentrados no afectó significativamente a ninguno de los parámetros asociados a la degradabilidad de la PB. Se destaca la elevada fracción soluble (52-60%) lo que resulta compatible con los elevados valores de degradabilidad efectiva observados para la PB (superiores al 80%; Cuadro 6) y las elevadas concentraciones de N-NH3 (Cuadro 4). La degradabilidad potencial de la PB resultó muy alta en todos los tratamientos, sin observarse diferencias estadísticamente significativas entre los mismos (T0 = 97%, T1 = 96.9% y T2 = 96.3%; P< 0.85). Producción y composición de la leche Los resultados de producción y composición de la leche se presentan en el Cuadro 8. El aporte de lípidos insaturados no afectó la producción de leche ni la concentración o producción de los principales componentes de la misma (Cuadro 8). Aunque el tenor graso de la leche sufrió una importante disminución (-5.9 gramos por kg comparado con T0) ante el aporte de lípidos insaturados, no pudo detectarse significancia estadística. Discusión El principal objetivo del presente experimento fue verificar que la suplementación con sales cálcicas de ácidos grasos insaturados no traía consecuencias digestivas negativas a nivel ruminal en vacas lecheras en pastoreo cuya dieta base estaba constituída por forraje fresco de alta calidad. Para mantener cierta constancia entre períodos experimentales en la calidad del forraje ofrecido se utilizó un sistema de pastoreo rotativo en franjas diarias manteniendo a su vez una asignación de forraje no inferior a 30 kg MS/vaca/día. El valor de asignación fijado y la biomasa promedio obtenida a lo largo del trabajo (2764 kg MS/ha) resultan ser condiciones predisponentes para maximizar el consumo de forraje (Minson, 1990). El sistema de pastoreo implementado pareció ser eficaz para mantener una alta calidad nutritiva en la pastura de alfalfa ya que la misma no experimentó grandes variaciones entre los diferentes períodos experimentales (Cuadro 2). Según Paterson et al. (1994) los máximos consumos en pasturas base alfalfa se obtendrían con contenidos de FDN y FDA menores al 42% y 30%, respectivamente, valores que resultan superiores a los observados en el presente ensayo (22-29% de FDN y 1720% de FDA). Además, ha sido sugerido (Wolter, 1994) que niveles de FDA del orden de 19-20% y de FDN del orden de 28-34% resultan ser los indicados a fines de obtener una adecuada fermentación ruminal en vacas lecheras de alto potencial de producción. El contenido de PB de la pastura estuvo cercano al valor máximo del rango (15 a 25% PB) propuesto por Minson (1990) a fines de obtener una alta digestibilidad ruminal de la MS. Estos resultados indicarían que tanto la cantidad de pastura ofrecida como su calidad fueron no limitantes para obtener elevados consumos de forraje y patrones normales de fermentación ruminal. En dichas condiciones, es posible realizar una correcta evaluación de los posibles efectos de los tratamientos experimentales sobre el ambiente y la digestión ruminal. El porcentaje de proteína de los concentrados (Cuadro 3) resultó máximo en T0 (14%) para experimentar ligeras reducciones ante la inclusión de AG-Ca. Este resultado se explica por el efecto de dilución de la proteína (T1) o reemplazo (T2) de proteína (maíz por lípidos) que los AG-Ca generan al ser incluidas. La digestibilidad de los concentrados disminuyó ante la inclusión de lípidos en la mezcla. Este resultado es consecuencia de la metodología in vitro utilizada (Tilley y Terry, 1963), la cual no está diseñada para evaluar la digestibilidad de alimentos que contienen una alta concentración de grasas. Efectos sobre el ambiente ruminal Ante un posible efecto negativo del aporte de lípidos no protegidos adecuadamente podría esperarse, junto con una menor digestión ruminal de la FDN, un aumento en las proporciones de ácido propiónico (C3), una disminución en la proporción de ácido acético (C2) y de ácido butírico y una menor concentración total de AGV (Jenkins, 1993). Además, las altas dosis de AG-Ca (0.8 kg/vaca/día) utilizadas en el presente trabajo serían suficientes para inducir modificaciones en el ambiente ruminal si no fueran completamente inertes. Sin embargo, la concentración de AGV totales no presentó diferencias significativas entre tratamientos, al igual que las proporciones de C3, C2, ni la relación C2:C3, tanto en condiciones de adición como de sustitución de almidón por lípidos en el concentrado (Cuadro 4). En el presente trabajo, el hecho de haber trabajado en condiciones de adición (T1) permitió evaluar los efectos puros de los lípidos aportados sobre el perfil de fermentación generado. La inexistencia de efectos a nivel ruminal sobre el patrón de fermentación generado en ambos tratamientos sugiere una adecuada protección (ausencia de disociación) de las AG-Ca, aún ante los bajos valores de pH registrados (Figura 1). El resultado es a su vez coherente con la ausencia de efectos de los AG-Ca sobre la degradación ruminal de la FDN (ver más adelante, Cuadro 5). Sólo la concentración de isobutirato presentó diferencias significativas entre tratamientos, siendo mayor en el tratamiento sin lípidos (T0, Cuadro 4). Dado que los isoácidos constituirían factores de crecimiento para las bacterias (Klusmeyer et al., 1987) podría postularse un mayor crecimiento bacteriano en el T0. Sin embargo, los niveles de N-NH3 en rumen no resultaron menores en el T0 (Cuadro 4). Puesto que no se realizaron mediciones de flujo de proteína bacteriana a duodeno, no resulta posible establecer si la mayor concentración de isobutirato en el T0 se debe a una menor utilización por parte de las bacterias (y por lo tanto una menor síntesis de proteína bacteriana) o si realmente hubo mayor síntesis de proteína microbiana. Además, DePeters y Cant (1992) postularon que el reemplazo de una parte del concentrado por grasa protegida en la dieta de rumiantes incrementaría la concentración de N-NH3 en rumen debido a un aumento de la relación proteína/energía fermentescible en rumen de la dieta, ya que los AG no son utilizados como fuente energética a nivel ruminal. Esto no fue observado en el T2 (sustitución), donde la concentración promedio de N-NH3 no difirió significativamente de las del resto de los tratamientos (Cuadro 4). Previos estudios conducidos en condiciones de pastoreo indicaron ausencia de efectos significativos del aporte de lípidos protegidos sobre la concentración de N-NH3 tanto en adición (Schroeder et al., 2002) como en sustitución (Salado et al., 2004). Por lo tanto, la ausencia de efectos de las AG-Ca sobre la proteólisis ruminal (Cuadro 7), los similares valores de N-NH3 (Cuadro 4) y de proteína láctea (Cuadro 8) sugerirían ausencia de efectos significativos de los lípidos sobre la síntesis de proteína microbiana, aún en el caso de reemplazo isoenergético de almidón por lípidos (T2). El elevado contenido de CNES de las pasturas (Cuadro 2) asi como la elevada digestibilidad de la FDN (Figura 2) pueden haber compensado el efecto del reemplazo de carbohidratos (almidón) por grasas en el tratamiento T2. El pH ruminal es generalmente poco afectado por la suplementación con lípidos (Palmquist y Jenkins, 1980; Tamminga y Doreau, 1991), observación que resultó consistente con los resultados del presente estudio. En efecto, la presencia de las AG-Ca en la ración no modificó los valores promedio de pH ruminal (Cuadro 4), resultado también observado en otros trabajos en condiciones de alimentación pastoril y suplementación con lípidos saturados (Schroeder et al., 2002; Salado et al., 2004) o en alimentación con raciones completamente mezcladas (Chan et al., 1997; Khorasani y Kennelly, 1998). Como se aprecia en la Figura 1, los valores de pH ruminal (para cada horario y tratamiento) se mantuvieron durante la mayor parte del día por debajo de lo considerado como óptimo para la digestión de la fibra (pH cercanos a 7; Satter et al., 1999). Sólo en las horas 0 y 12 se registraron mediciones superiores a 6, momentos en los cuales las vacas eran ordeñadas y se les suministraba el concentrado. Estos bajos valores de pH podrían explicarse por una elevada producción de AGV a nivel ruminal (Cuadro 4), a causa de las elevadas concentraciones de CNES (Cuadro 2) y la alta digestión de la FDN (Figura 2) de la pastura suministrada, y coinciden con los valores obtenidos en ensayos sobre pasturas de alta calidad (Schroeder et al., 2002; Schroeder et al., 2003; Salado et al., 2004). Según Satter et al. (1999) los microorganismos celulolíticos serían capaces de mantener estable su pH intracelular aún en un medio ácido, lo que explicaría que aún en rangos subóptimos de pH se obtengan adecuados niveles de digestión ruminal del forraje. Desaparición ruminal de la MS, FDN y PB del forraje La fracción soluble de la MS del forraje incubado tendió a ser mayor (P<0.06) en el tratamiento con sustitución isoenergética del grano de maíz por AG-Ca (T2) (Cuadro 5). Este resultado es difícil de explicar, ya que efectos sobre la fracción soluble no son de esperar ante la aplicación de tratamientos nutricionales como la suplementación con lípidos. Por otra parte, este resultado no modificó significativamente a la degradabilidad efectiva observadas en el T2, al compararlas con los demás tratamientos (Cuadro 5). El resto de los parámetros analizados no fue significativamente afectado por la inclusión de lípidos insaturados, tanto en condiciones de adición como de sustitución de grano de maíz por lípidos (Cuadro 5). Este resultado resulta consistente con otros hallados tanto en experiencias en pastoreo (Schroeder et al., 2002; Schroeder et al., 2003; Salado et al., 2004) como utilizando raciones completamente mezcladas (Chan et al., 1997; Khorasani y Kennelly, 1998). Los altos porcentajes observados de MS soluble (45-50%) confirman la alta calidad de la pastura de alfalfa utilizada (Cuadro 2) y coinciden con los hallados por otros autores en similares condiciones (Elizalde et al., 1999). La tasa de degradación de la MS (9-11%/h) tampoco fue afectada por los tratamientos y resultó menor a la informada (13.8%/hora) por Elizalde et al. (1999) para pasturas de alta calidad. El efecto depresivo de los lípidos sobre la actividad celulolítica a nivel ruminal resulta ser más frecuente con los AG insaturados que con los AG saturados (Jenkins, 1993). Además, los niveles de pH ruminal obtenidos en nuestro estudio (5.8) fueron cercanos al pH (5.6) al cual las sales cálcicas de AG insaturados podrían disociarse significativamente (Sukhija y Palmquist, 1990). Los AG insaturados libres en rumen podrían afectar negativamente a la población de bacterias con actividades celulolíticas (Tamminga y Doreau, 1991; Jenkins, 1993; Doreau y Ferlay, 1994) y asi reducir la digestión de la FDN. Garciarena et al. (1990) observaron una tendencia a la disminución de la tasa de degradación de la FDN del forraje (avena) ante la inclusión por sustitución de 500 g diarios de AG-Ca a vacas lecheras en pastoreo, efecto que sería explicado por una inadecuada protección de los lípidos utilizados. En el presente ensayo, sin embargo, no se detectaron diferencias significativas en ninguno de los parámetros asociados a la degradación de la FDN (Cuadro 5). La ausencia de un tiempo de retardo en la desaparición de FDN fue coincidente con lo encontrado por otros autores evaluando forrajes frescos de alta calidad (Elizalde, 1999; Schroeder et al., 2002) y concuerda con la elevada calidad del forraje consumido y las adecuadas condiciones ruminales para su degradación. Estos resultados, junto a la ausencia de efectos del aporte de los lípidos sobre las variables asociadas al ambiente ruminal (Cuadro 4), sugieren que la saponificación de AG insaturados y su formulación como AG-Ca resultó una herramienta válida para evitar una posible interacción negativa con el metabolismo ruminal. Los parámetros asociados a la degradabilidad proteica del forraje incubado in situ no fueron afectados por los tratamientos (Cuadro 7), resultado coincidente con lo informado por otros autores trabajando en condiciones de pastoreo (Garciarena et al., 1990; Salado et al., 2004). Estos resultados son a su vez compatibles con la ausencia de efecto de los lípidos sobre las concentraciones ruminales de N-NH3 (Cuadro 4). Producción y composición de la leche Los efectos de la suplementación con lípidos sobre la producción de leche parece ser altamente dependiente del grado de saturación de los AG utilizados (Gagliostro y Chilliard, 1992; Schroeder et al., 2004). Según una reciente revisión bibliográfica (Schroeder et al., 2004), mientras que la suplementación con AG saturados consistentemente ocasiona incrementos en la producción de leche en vacas lecheras en pastoreo, dichos efectos parecen ser de menor magnitud y mas variables cuando son suplementados AG insaturados. En el presente ensayo, ni la producción de leche ni la concentración de los componentes de la misma se vieron significativamente afectados por el aporte de lípidos insaturados (Cuadro 8). Estos resultados son fundamentalmente descriptivos ya que la duración de los períodos experimentales puede considerarse insuficiente como para detectar efectos significativos de la suplementación implementada. Aunque estadísticamente no fue significativo, el contenido graso de la leche disminuyó marcadamente (-5.9 gramos por kg de leche) debido a la supplementación con AG-Ca (Cuadro 8). Este resultado es coincidente con lo encontrado en la mayoría de los trabajos con suplementación con AG insaturados a vacas lecheras consumiendo pasturas de alta calidad (Schroeder et al., 2004). Se ha postulado que la disminución del tenor graso de la leche ante el aporte de AG insaturados sería consecuencia de una acción directa de los AG sobre la síntesis grasa a nivel de la glándula mamaria (Bauman y Griinari, 2001; Schroeder et al., 2004). Los resultados del presente trabajo parecen apoyar dicha hipótesis ya que la reducción en el tenor graso de la leche no estuvo asociada a efectos negativos sobre la digestión ruminal de la fibra del forraje. Sin embargo, es necesaria más investigación a fin de evaluar los efectos a más largo plazo de la suplementación con este tipo de AG-Ca sobre la producción y composición de la leche en vacas lecheras en pastoreo. Conclusiones En vacas lecheras en condiciones de alimentación pastoril, la inclusión de ácidos grasos insaturados bajo la forma de sales cálcicas, tanto en condiciones de adición como de sustitución isoenergética, no afectó a los parámetros de ambiente ruminal ni la degradación ruminal del forraje. Estos resultados sugieren que los AG insaturados y saponificados pueden constituir una fuente de energía apta para la alimentación de rumiantes sin perturbar la digestión ruminal, aún en condiciones de alimentación pastoril con bajos valores de pH ruminal como los observados en el presente experimento. Literatura Citada
© 2007 ALPA. Arch. Latinoam. Prod. Anim. The following images related to this document are available:Photo images[la07014t4.jpg] [la07014f1.jpg] [la07014t5.jpg] [la07014t2.jpg] [la07014t1.jpg] [la07014f2.jpg] [la07014t8.jpg] [la07014t6.jpg] [la07014t7.jpg] [la07014t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}