 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
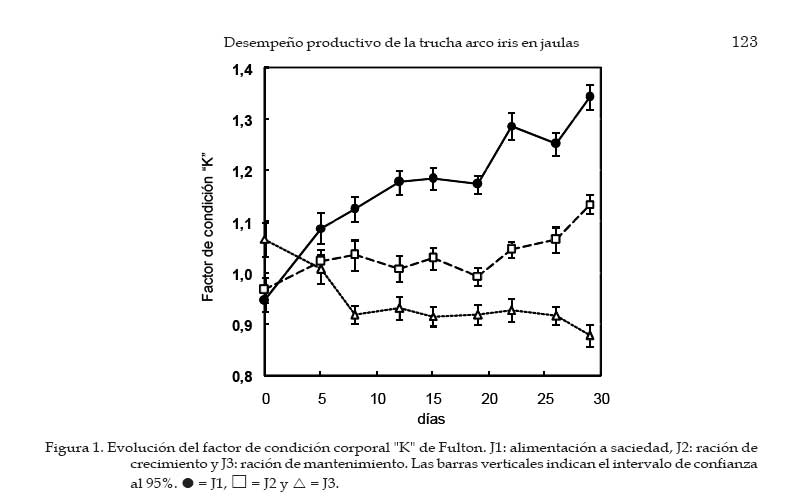
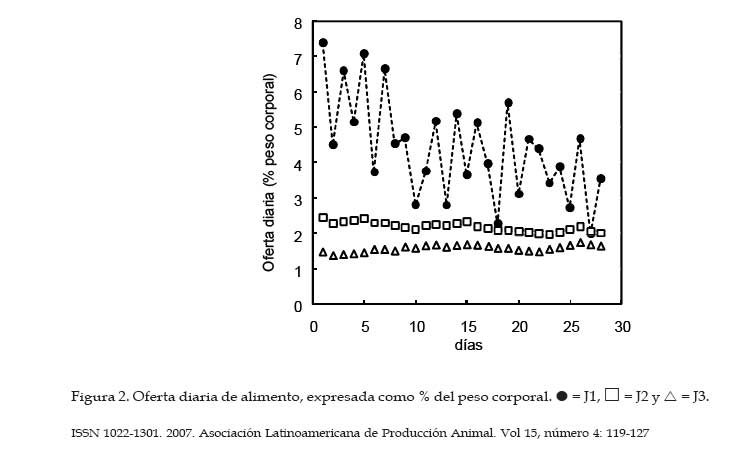
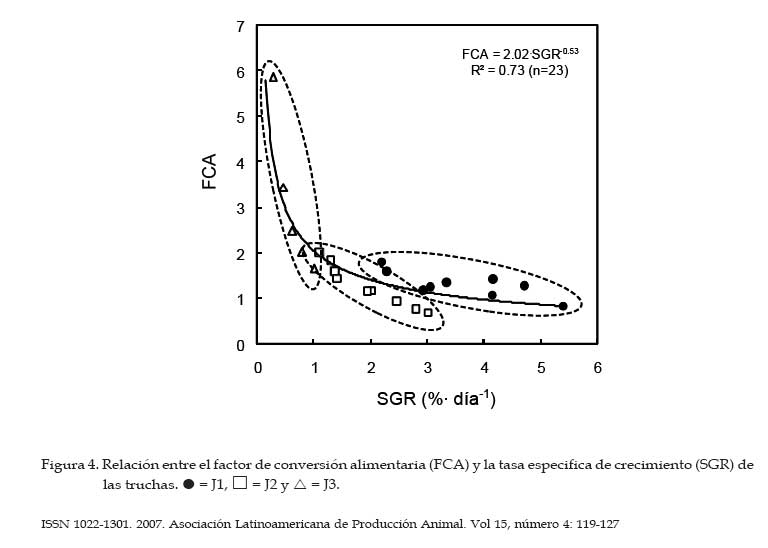
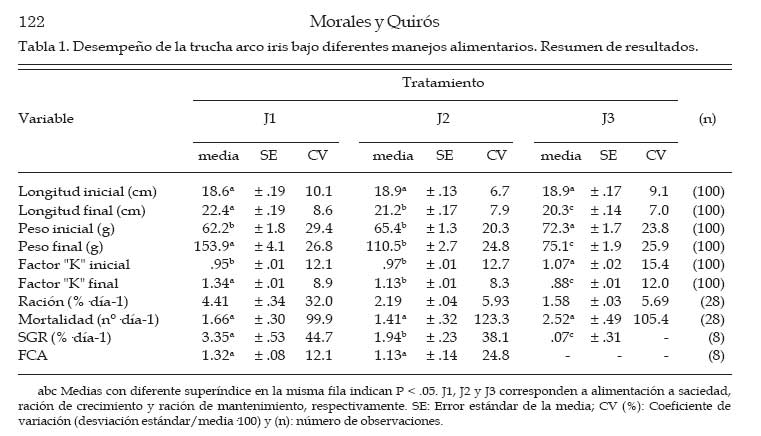
Archivos Latinoamericanos de Producción Animal, Vol. 15, No. 4, 2007, pp. 119-127 Desempeño productivo de la trucha arco iris en jaulasbajo diferentes estrategias de alimentación Productive performance of rainbow trout in cages underdifferent feeding strategies G.A. Morales y R. Quirós Área de Sistemas de Producción Acuática, Departamento de Producción Animal, Facultad de Agronomía, Universidad de Buenos Aires. Buenos Aires, Argentina. Recibido Mayo 15, 2006. Aceptado Agosto 16, 2007 Code Number: la07018 ABSTRACT Slight improvements in the feeding efficiency can be translated in an important diminution of the production costs and feed wastes that would alter the water quality. With the objective of determine the optimal daily ration during the season of greater growth and study the effect of ration on the corporal condition (K) of rainbow trout under the conditions of a commercial operation, the growth of 15 000 juveniles under 3 treatments was evaluated: feeding to satiety (J1), growth ration (J2) and maintenance ration (J3). The total length, corporal weight and K differed significantly with the applied treatment, resulting in all cases J1>J2>J3. At the final of trial, J1 group has shown the higher body weight (153.9 ± 4.12 g) with respect to J2 (110.5 ± 2.74 g) and J3 (75.1 ± 1.94 g). An oscillating feeding behavior, with variations from 1 to 3% of a day for another one, has been observed in J1. The relation between the specific growth rate (SGR) and the daily ration (R) fit to a quadratic function, SGR=(-0.22.R2)+(2.57.R)-3.12. On the other hand, the relation between the feed conversion ratio FCA and the SGR fit to the function FCA=2.02.SGR-0.53. Optimal values to FCA have been observed when fed at 50% of satiety or more. No significant differences were observed for global FCA between J1 (1.32 ± 0.08) and J2 (1.13. ± 0.14). Consequently, satiety feeding would be the best option during the summer when the temperature is optimal for the growth. Key words: Oncorhynchus mykiss, Cage aquaculture, Specific growth rate, Feed conversion ratio, Corporal condition. RESUMEN Leves mejoras en la eficiencia alimentaria pueden traducirse en una importante disminución de los costos de producción y de desechos de alimento que alterarían la calidad del agua. Con el objetivo de determinar la ración diaria óptima durante la estación de mayor crecimiento y estudiar el efecto del tamaño de ración sobre la condición corporal (K) de la trucha arco iris bajo las condiciones de un criadero comercial, se evaluó el crecimiento de 15 000 juveniles bajo 3 tratamientos: alimentación a saciedad (J1), ración de crecimiento (J2) y una ración de mantenimiento (J3). La longitud total, el peso corporal y el K difirieron significativamente con el tratamiento aplicado, resultando en todos los casos J1>J2>J3. Al final de la experiencia, J1 alcanzó el mayor peso corporal (153.9 ± 4.12 g) respecto de J2 (110.5 ± 2.74 g) y J3 (75.1 ± 1.94 g). J1 mostró un comportamiento alimentario oscilante, arrojando variaciones de 1 a 3% de un día para otro. La relación entre la tasa de crecimiento específico (SGR) y la ración diaria (R) ajustó a la función SGR=(-0.22.R2)+(2.57.R)-3.12. Por otro lado, la relación entre el factor de conversión del alimento (FCA) y el SGR ajustó a la función FCA=2.02.SGR-0.53. Valores óptimos de FCA se observaron con raciones al 50% de saciedad o mayores. No se encontraron diferencias significativas para FCA global entre J1 (1.32 ± 0.08) y J2 (1.13. ± 0.14). Consecuentemente, alimentar a saciedad sería la mejor opción durante el verano cuando la temperatura es óptima para el crecimiento. Palabras clave: Oncorhynchus mykiss, Cultivo en jaulas, Tasa de crecimiento específico, Factor de conversión alimentaria, Condición corporal. Introducción La alimentación generalmente representa el mayor costo operativo de una explotación intensiva de engorde de peces (Huguenin y Ansuini, 1978; Chua y Teng, 1980; Kim, 1981; Sungkasem, 1982) y según Higgs et al. (1995) pueden constituir aproximadamente el 55% de los costos de producción anuales totales de los criaderos comerciales. Elevados costos de producción asociados a la alimentación frecuentemente se relacionan a una pobre eficiencia en la conversión alimentaria, lo que resulta además en una importante descarga de nutrientes al medio acuático (Costello, et al., 1996). Un objetivo primario de los productores es engordar peces tan rápida y eficientemente como sea posible. Sin embargo, varios estudios (Elliot, 1976; Brett y Groves, 1979; Storebakken y Austreng, 1987b; Kolsater, 1995; Wendall, 2004) indican que si bien para alcanzar el máximo crecimiento se debe alimentar ad-libitum o "a saciedad", cuando se utiliza dicho manejo alimentario disminuye la eficiencia de conversión del alimento. Una herramienta muy utilizada para determinar la ración diaria a suministrar consiste en utilizar tablas de alimentación difundidas por las empresas que fabrican alimento. Sin embargo, estas suelen estar basadas en modelos teóricos basados en la temperatura del agua y tamaño de los peces (Haskell, 1959 modificado por Westers, 1995; Leitritz y Lewis ,1980; Iwama y Tautz, 1981; From y Rasmussen, 1984; Smith, 1989) que deben ser ajustados a las condiciones específicas de producción (Cho y Boreau, 1997) y pueden sobrestiman la cantidad de alimento a suministrar (Cerdá, 2000). Varios autores (Dabrowski, 1984; Uys y Hetcht, 1985; Charlon y Bergot, 1986) han propuesto que las pérdidas de eficiencia alimentaria, han sido a menudo el resultado de un exceso o falta de alimentación, probablemente, a causa del uso de tablas de alimentación no adaptadas a las condiciones de locales de cultivo. El objetivo de este trabajo fue determinar la ración óptima de alimento a suministrar diariamente a juveniles de trucha arco iris Oncorhynchus mykiss (Walbaum, 1792) durante la estación de mayor crecimiento, asimismo evaluar el efecto de la estrategia alimentaria utilizada sobre la condición corporal de las truchas bajo las condiciones de un criadero comercial. Materiales y Métodos Instalaciones El estudio se llevó a cabo en el criadero comercial Truchas Alicurá S.R.L. situado en el embalse Alicurá, noroeste de la Patagonia Argentina (40º41´44´´S 71º01'01''O). Se utilizaron 3 jaulas de 50 m3 con mallas de calibre de tejido de 1.4 cm, las cuales fueron acondicionadas, lavadas y desinfectadas con solución de yodo 10% (vol/vol) antes de su uso. Sobre la superficie de las jaulas se colocaron redes de protección (pajareras) con el fin de evitar la posible predación por parte de las aves. Manejo de peces La clasificación y conteo de 45 000 juveniles de trucha arco iris (65 ± 7 g de peso corporal) se realizó con un equipo de selección mecánica y conteo electrónico (Bioscanner, V-Channel 1). Fueron separados tres sub-lotes: cabeza, núcleo y cola de lote. El núcleo central de 15 000 individuos, fue dividido aleatoriamente en tres grupos experimentales de 5 000 individuos cada uno. El suministro de alimento se llevó a cabo en forma manual y controlada. La entrega de alimento se efectuó en dos raciones diarias: por la mañana entre las 08:00 y las 09:00 h y por la tarde entre las 16:00 y las 17:00 h. Se utilizó un alimento balanceado extruído para truchas con calibre de partícula de 3 mm (GANAVE), con una composición centesimal de 46% PB, 17% EE, 2% FB, 18% CEN, 90% MS (base MH) y 2 700 Kcal/kg de EM (base MS). Las variables físico-químicas registradas fueron la temperatura media diaria del agua (TMD) y la concentración de oxígeno disuelto en el agua (OD), utilizando para ello una sonda limnológica (Lutron, DO-5510). Las mediciones fueron efectuadas tres veces al día (9, 12 y 17 h) a una profundidad constante de un metro. Las muestras de peces se tomaron (24 h de ayuno) de diferentes sitios y profundidades de la jaula. Los peces fueron anestesiados con una solución acuosa de benzocaína (1:10 000. Una vez anestesiados, se registraron la longitud total y el peso de cada pez. La longitud se midió utilizando un ictiómetro (+/- 0.5 mm), mientras que el peso corporal de cada pez se registró por medio de una balanza portátil Acculab V-1200 (+/- 0.1 g). Cálculo de raciones La ración suministrada a cada uno de los tratamientos se calculó utilizando el modelo de crecimiento de Haskell (1959) modificado por Westers (1995). La función utilizada para la determinación de la ración de mantenimiento fue: Rm = 2·TMD·(100·K·L) 1. Donde Rm (en % del peso corporal·día-1); 2: Constante de Haskell; TMD: Temperatura media diaria del agua (ºC); K: Factor de Condición Corporal de "Fulton"; L: Longitud total del pez (cm). Para el cálculo de la ración de crecimiento se utilizó el modelo mencionado anteriormente afectado por una constante de crecimiento (C), la cual indica la proporción de la energía que es canalizada a crecimiento. Con la ración de mantenimiento se pretende que la trucha no cambie significativamente de peso. Sin embargo, con la ración de crecimiento se intenta generar un excedente de energía que sería destinado a crecimiento (energía retenida) y corresponde al 30% de la energía bruta ingerida en los salmónidos (Kaushik y Médale, 1994). La función utilizada para determinar la ración de crecimiento (Rc) fue: Rc = (2·TMD·(100·K·L)-1) / (1 - C). Durante el suministro de alimento a saciedad, el momento en el cual los peces alcanzaban la saciedad se determinó en base al comportamiento alimentario de los mismos. Poca actividad en superficie, natación profunda y en cardumen, una baja actividad predadora y alimento sin ser consumido por debajo 50 cm de profundidad aún con bajas frecuencias de alimentación, fueron los indicadores visuales que se utilizaron para determinar el momento en que se detenía el suministro de alimento. Diseño experimental y análisis estadístico Se utilizaron tres grupos experimentales de 5 000 individuos cada uno, tomando como unidad experimental el grupo de animales que corresponde a un determinado tratamiento. A cada grupo se le asignó en forma aleatoria una jaula y un tratamiento, a los que se les aplicaron distintas raciones de alimento: alimentación a saciedad (J1), ración de crecimiento (J2) y ración de mantenimiento (J3). Durante 28 días de experiencia (mes de febrero) se llevaron a cabo nueve muestreos, capturando 100 individuos (n=100) cada vez en forma aleatoria a distintas profundidades de la jaula. Los mismos se realizaron durante el primer y cuarto día de cada semana. Una vez anestesiados, se registró el peso vivo y la longitud total individual. También se registró la mortalidad diaria. Las variables biológicas calculadas fueron la tasa de crecimiento específico (SGR), el factor de conversión alimentaria (FCA) y el factor de condición corporal de "Fulton" (K): SGR = [(ln(peso final)-ln(peso inicial))/días]·100 FCA = (alimento entregado)/(ganancia de peso vivo) K (Fulton) = [peso individual/(longitud total)3]·100 Los datos se procesaron mediante un análisis de varianza (ANOVA) para medidas repetidas (Sokal y Rohlf, 1995). Para el análisis se tuvieron en cuenta los efectos del factor tratamiento (entre-sujetos) y tiempo (intra-sujetos), la interacción entre ambos y se utilizó como co-variable el peso inicial y la condición corporal. La detección del momento en que las diferencias entre medias de los tratamientos comenzaron a ser significativas (P<.05) se determinó por medio del test de Tuckey HSD. Se utilizó la media aritmética como estimador de tendencia central y el error estándar (SE) como medida de dispersión, previa constatación de normalidad distributiva (Wilk-Shapiro). El coeficiente de variación (CV) de las medias obtenidas se calculó como la desviación estándar/media·100. Se estableció un valor P<.05, por debajo del cual se rechazaría la hipótesis nula de igualdad poblacional. Las variables SGR, FCA y R se relacionaron por medio de un análisis de regresión, presentando la función a la que se ajustan los datos, el número de observaciones (n) y su coeficiente de determinación (R2). Todos los análisis estadísticos se realizaron con el programa Number Cruncher Statistical Systems NCSS 2000. Resultados Al cabo de 4 semanas de tratamiento los grupos J1 y J2 tuvieron un incremento de biomasa respecto de la inicial de 145% y 67%, respectivamente, en tanto J3 (0.02%) mantuvo su biomasa inicial. La densidad inicial fue de (6.2 ± 0.3 kg/m3) para los tres grupos experimentales, difiriendo la densidad final con el tratamiento aplicado (J1= 15.2 kg/m3, J2= 11 kg/m3; J3= 7.4 kg/m3). Los valores registrados de mortalidad diaria media (1.66, 1.41 y 2.52 individuos·día-1 para J1, J2 y J3, respectivamente) fueron normales para las condiciones del ensayo, no encontrando diferencias significativas entre los grupos experimentales (P = .12). Las mediciones de oxígeno disuelto en el agua registradas tuvieron una media de 11.4 ± 0.19 mg/l OD y no se registraron valores por debajo de 8.9 mg/l OD. La temperatura media del agua fue de 14.9 ± 0.1ºC, registrando un valor mínimo de 13.9ºC y máximo de 16.4ºC. El análisis de varianza de medidas repetidas arrojó diferencias significativas (P<.05) para longitud total, el peso corporal y el factor de condición corporal de los peces (Cuadro 1), resultando en todos los casos J1 > J2 > J3 al final de la experiencia. El peso corporal de los peces mostró diferencias significativas en distintos momentos desde el inicio de la experiencia. A los 8 días de tratamiento se diferenciaron J1 de J3, a los 12 días J1 de J2 y a los 15 días J2 de J3. Al final de la experiencia J1 alcanzó un peso corporal de 153.9 ± 4.12 g (CV= 26.8%), seguido por J2 (110.5 ± 2.74 g; CV= 24.8%) y J3 (75.1 ± 1.94 g; CV= 25.9%). Este último, prácticamente no mostró cambio en el peso corporal de los peces (3,9%), aunque evidenció un mayor incremento relativo en la longitud total de los peces (7,4%) lo que se tradujo en una importante disminución en la condición corporal de los peces. El K inicial difirió significativamente entre J1 y J3 y entre J2 y J3 (P < .0001), no así entre J1 y J2. A pesar de ello, J3 disminuyó rápidamente su condición corporal, mostrando una tendencia decreciente durante el transcurso de la experiencia, J2 mantuvo su K con una leve mejora hacia el final y J1 mostró una constante mejora hasta el final (Figura 1). De la misma manera que para peso corporal, a los 8 días desde el inicio se encontraron diferencias significativas para K entre J1 y J2, mientras que J1 y J3 como J2 y J3 se diferenciaron a los 12 y15 días, respectivamente. El tratamiento J1 arrojó el mayor K final (1.34 ± 0.12), seguido por J2 (1.13 ± 0.09) y J3 (0.88 ± 0.10), respectivamente. El tratamiento J1 entregó la mayor tasa de crecimiento específica (SGR= 3.35 ± 0.53%·día-1), seguido por J2 (1.94 ± 0.23%·día-1). Sin embargo, el SGR del tratamiento J3 (ración de mantenimiento) prácticamente fue nulo (0,07 ± 0.31%·día-1). La ración diaria media asignada a cada tratamiento correspondió a 4.41 ± 0.34 (J1), 2.19 ± 0.04 (J2) y 1.6 ± 0.03%·día-1 (J3). En la Figura 2 se puede observar una marcada diferencia en la dispersión de estos valores medios. El tratamiento J1 evidenció un comportamiento alimentario oscilante, arrojando variaciones en la demanda de alimento de 1 a 3 %·día-1 de un día para otro (CV= 32.0%). En tanto, la dispersión de los valores de ración diaria suministrada a J2 (CV= 5.93%) y J3 (CV= 5.69%), basadas en el modelo de la TMD y el tamaño del pez, fue sustancialmente menor. La relación entre el SGR y la R suministrada (Figura 3) fue curvilínea, ajustando a un modelo de regresión polinomial de segundo orden, SGR = (-0.22.R2) + (2.57.R) - 3.12; (R2= 0.75; n= 27). Se observó que una R igual a 1.3%·día-1 del peso corporal equivale a un SGR nulo. A mayores niveles de oferta de alimento, el incremento en el SGR por unidad de ración suministrada fue cada vez menor, disminuyendo la pendiente de la curva. Los puntos cercanos a un SGR= 0 correspondieron al tratamiento J3, mientras que los valores registrados para J1 se encontraron en la zona de mayor ración suministrada y menor pendiente. Por cierto, los valores de J2 se encontraron agrupados en la zona de pendientes medias entre J3 y J1. Al relacionar el FCA con el SGR (Figura 4), se observó que a medida que los peces experimentan mayores tasas de crecimiento, la eficiencia de conversión mejora. Valores de SGR menores a 1.0%·día-1 se relacionan con un elevado FCA (aprox. de 2.0 a 6.0), esta baja eficiencia alimentaria corresponde al los valores registrados para el tratamiento J3. Valores de SGR entre 1.0 y 5.5%·día-1, correspondientes a J1 y J2, evidenciaron una mejor conversión alimentaria (aprox. de 1.0 a 2.0). Al comparar el FCA global (alimento total suministrado / incremento de la biomasa total), donde J1 (1.32 ± 0.08) fue levemente mayor a J2 (1.13. ± 0.14), estos no difirieron estadísticamente (P = .91). La relación entre el FCA y el SGR (Figura 4) ajustó a la función potencial FCA = 2.02.SGR-0.53 (R2= 0.73; n= 23). Se presentaron sólo los valores positivos del tratamiento J3, ya que durante el transcurso del ensayo las truchas bajo tratamiento de mantenimiento tuvieron pequeñas pérdidas de peso. Discusión y Conclusiones Los cambios observados en la condición corporal de los peces según el tratamiento aplicado demuestran que el uso de una estrategia de producción basada en restricciones alimentarias frecuentes, hasta el nivel e mantenimiento, genera una importante pérdida de condición corporal de las truchas. Este aspecto se puede observar en los individuos del grupo J3, los cuales no aumentaron significativamente su peso corporal, pero sí su longitud. Si bien el "frenar" lotes en producción por restricción alimentaria es una herramienta de manejo comúnmente utilizada en condiciones de escasez de alimento, baja demanda del producto en el mercado o simplemente cuando se desea generar un gradiente de cosecha a lo largo del año, no sería recomendable realizar dicho manejo por periodos prolongados. En el caso de que el productor necesite utilizar provisoriamente una estrategia alimentaria basada en una ración de mantenimiento (J3), de acuerdo a la función obtenida que relación entre el SGR y la ración diaria suministrada (Figura 3), ésta no debería encontrarse por debajo de 1.3%·día-1 del peso corporal cuando la temperatura del agua es óptima para el crecimiento. Una ración menor a dicho valor generaría no solo una disminución en la condición corporal de las truchas, sino que los peces perderían peso y por tanto disminuiría la biomasa de los lotes en producción afectados a dicha estrategia alimentaria. Los registros de consumo de alimento evidenciaron una gran variación en el apetito de las truchas. Este comportamiento alimentario oscilante observado en el tratamiento con alimentación a saciedad ha sido descrito previamente para salmónidos en varios estudios (Grove et al., 1978; Juell et al., 1993, 1994; Bailey y Alanärä, 2001). Estas variaciones diarias en el consumo de alimento estarían relacionadas con el efecto residual que genera el elevado consumo de alimento sobre el llenado del tracto digestivo y el tiempo requerido para su depleción (Grove et al. 1978; Bromley, 1994). Una elevada ingestión de alimento afectaría las señales periféricas del aparato digestivo involucradas en la generación de apetito, resultando en un menor consumo al día siguiente, para luego, a las 48-72 h recuperar el apetito "normal" y volver a ingerir mayor cantidad de alimento. En el presente estudio el factor de conversión alimentaria (FCA) global no difirió significativamente entre el tratamiento alimentación a saciedad (J1) y la ración de crecimiento (J2). A partir de la función que relaciona el FCA y la tasa de crecimiento de los peces (SGR), se observó que al trabajar con una baja tasa de crecimiento, el factor de conversión es elevado, es decir, la eficiencia alimentaria es baja. A medida que la tasa de crecimiento aumenta, el factor de conversión disminuye, mejorando la eficiencia alimentaria hasta alcanzar un óptimo biológico, el cual se mantiene hasta los valores máximos de crecimiento logrados con una alimentación a saciedad (máximo consumo voluntario de alimento). Estos resultados, obtenidos bajo las condiciones de infraestructura y manejo de un criadero comercial, concuerdan con los obtenidos por Alanärä (1994), Bureau (1997) y Azevedo (1998). En este último estudio se evaluó el efecto de tres raciones (cercano a saciedad, 85% y 70% de saciedad) sobre el crecimiento de la trucha arco iris a diferentes temperaturas. Los autores concluyeron que la eficiencia de conversión alimentaria no fue afectada por los diferentes niveles de alimentación utilizados. Por otro lado, Storebakken y Austreng (1987a) en un estudio realizado con salmón del atlántico reportó que no hubo diferencias en la eficiencia alimentaria en peces alimentados a niveles de 50, 75 o 100% de la ración requerida para obtener el máximo crecimiento. Los resultados obtenidos en el presente estudio también concuerdan con Storebakken y Austreng (1987a), ya que los valores de ración diaria media entregada indican que J2 (2.19%·día-1) correspondió al 50% de J1 (4.41%·día-1) la ración requerida para obtener el máximo crecimiento, sin detectarse diferencias significativas en la eficiencia de conversión alimentaria. Sin embargo, estos resultados se contradicen con los publicados por varios autores (Elliot, 1976; Brett y Groves, 1979; Storebakken y Austreng, 1987b; Kolsater, 1995) que indican que la mejor eficacia alimentaria se presenta solamente a niveles de alimentación menores a los requeridos para obtener el máximo crecimiento, disminuyendo la eficiencia alimentaria cuando se trabaja a valores cercanos a la saciedad. Al comparar la ración diaria recomendada por otros autores se puede observar una gran variabilidad en los valores publicados, así Leitritz y Lewis (1980) y Smith (1989) indicaron que la ración diaria óptima para las condiciones de temperatura y peso corporal del presente estudio corresponden a 1.5 y 2.2%·día-1, respectivamente. En tanto al revisar los valores difundidos por algunas empresas elaboradoras de alimento balanceado para truchas (Alitec: 2.6; Ewos: 2.4; Skretting: 2.4%·día-1) se pueden observar distintos valores y con una tendencia mayor a los publicados por Leitritz y Lewis (1980) y Smith (1989). Ello apoyaría la hipótesis realizada por Cerdá (2000), quien sostiene que las tablas de ali-mentación difundidas por empresas elaboradoras de alimento balanceado sobrestiman la cantidad de alimento a suministrar. Sin embargo, debido a las diferencias en las tecnologías de fabricación (Zongjia y Hardy, 2003), la relación proteína/energía (Azevedo et al., 2004) y la digestibilidad (Bureau, et al., 1999) de los distintos alimentos para acuicultura, es difícil establecer una ración óptima para un determinado peso corporal y temperatura del agua. Por tanto, se hace necesario que el productor utilice las distintas tablas y modelos disponibles para el cálculo de raciones como una guía, para luego elaborar su propia base de datos con la cual pueda adaptar dichos modelos a las condiciones particulares de su criadero. De acuerdo a los resultados obtenidos y bajo las condiciones de cultivo del presente estudio, se concluye que si el productor desea obtener un rápido crecimiento de los peces, alimentar a saciedad sería la mejor opción durante el verano cuando la temperatura del agua se encuentra cercana al óptimo de la especie, siempre y cuando sea cuidadoso al entregar el alimento de manera que no se produzcan derroches por los lados o fondo de la jaula de cultivo. Literatura Citada
© 2007 ALPA. Archivos Latinoamericanos de Producción Animal The following images related to this document are available:Photo images[la07018f4.jpg] [la07018f3.jpg] [la07018f1.jpg] [la07018t1.jpg] [la07018f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}